| |
Методы психогенетики
Методы психогенетики
Основной целью психогенетических исследований является объяснение происхождения индивидуальных различий в психических и психофизиологических характеристиках человека. Для этого, во-первых, необходимо выяснить, какой вклад в изменчивость вносят наследственные и средовые факторы, и, во-вторых, по возможности попытаться объяснить механизмы влияния наследственности и среды на изучаемые характеристики.
Какими методами пользуется современная психогенетика для ответа на эти вопросы? До недавнего времени в распоряжении психогенетики имелись лишь генетико-эпидемиологические подходы, позволяющие работать на уровне популяций. При этом возможности для поиска механизмов наследственности были весьма ограниченными. Сейчас в связи со стремительным прогрессом молекулярно-генетических технологий генетика человека, и психогенетика в том числе, обогатились множеством новых методических приемов, открывающих широкие перспективы для проникновения в природу наследственных механизмов, участвующих в формировании психических особенностей человека.
в начало
Семейное и генетическое сходство
Одним из основных методических приемов генетики является семейное сравнение, т.е. сравнение организмов, объединенных родством. Г. Мендель, проводя опыты с горохом, изучал поколения родителей и потомков. Ф. Гальтон, анализируя родословные знаменитостей, сравнивал людей, связанных родственными узами. Из предыдущего изложения понятно, что основные экспериментальные подходы психогенетики также связаны с изучением различных категорий родственников. Прежде чем начать знакомство с конкретными экспериментальными методами психогенетики, рассмотрим, что лежит в основе сходства и различий между родственниками, и остановимся на некоторых особенностях измерения сходства.
Если мы предполагаем, что в основе изменчивости какого-либо признака лежит генетический компонент, то можно ожидать, что родственники будут более похожи по данному признаку, поскольку у них, скорее всего, имеются какие-то одинаковые гены, унаследованные от общего предка. У членов одной и той же семьи, помимо общих генов, как правило, имеются и общие средовые условия. Если признак небезразличен к влиянию среды, то на сходство родственников по этому признаку будут оказывать влияние не только общие гены, но и общая среда. Таким образом, семейное сходство включает в себя наследственный и средовой компоненты. Это создает определенные методические трудности в определении роли генов в формировании семейного сходства. Когда мы имеем дело с растениями или экспериментальными животными, мы легко можем нейтрализовать те воздействия, которые приводят к средовому сходству, поскольку имеем возможность выращивать потомство в любых необходимых условиях среды. Но мы не можем так поступить с семьями людей. Поэтому на формирование сходства между родителями и детьми, братьями и сестрами, близнецами и т.д. неизбежно будет влиять общая среда.
Необходимо различать сходство семейное и сходство генетическое. Многие черты являются семейными, не будучи наследственными. Например, в религиозных семьях чаще всего и родители, и дети исповедуют определенную религию, посещают храм и совершают религиозные обряды, но вряд ли кто решится утверждать, что такого рода поведение наследуемо, поскольку наблюдается у членов одной семьи. Это типичный случай семейного, но не генетического сходства.
Классическим стал пример семейного сходства, которое долгое время считалось генетическим, но при более детальном рассмотрении оказалось чисто средовым - это болезнь куру. Она представляет собой прогрессирующее нервное расстройство со смертельным исходом. До недавнего времени заболевание часто встречалось в одной из областей Новой Гвинеи. Болезнью чаще всего поражались члены одной семьи, причем основную часть пораженных составляли женщины. Данные статистики заболевания вполне соответствовали модели наследственной болезни с половыми различиями в экспрессивности (т.е. различной степени выраженности действия гена) у гетерозигот. Позднее выяснилось, что эта болезнь вызывается вирусом, поражающим мозг, а семейный характер заболевания объяснялся существованием в этих племенах традиций каннибализма: родственники умершего человека съедали кусочки его мозга и таким образом заражались вирусом куру. После введения законов, запрещающих каннибализм, заболевание было почти полностью ликвидировано.
Семейное сходство очень часто интерпретируют как наследственное, не имея к тому достаточных оснований. Причиной этого являются предвзятые социальные установки. Например, семейное сходство по степени религиозности или политическим убеждениям обычно не считается наследственной чертой, но вместе с тем многие уверены, что сходство между родителями и детьми в музыкальных способностях является следствием влияния общих генов, хотя в последнем случае, в принципе, возможно и иное, чисто средовое, объяснение. Как правило, в музыкальных семьях дети воспитываются в определенных традициях, которые предполагают раннее приобщение детей к музыке и стимулируют развитие музыкальности.
Таким образом, семейное сходство - это лишь то, что мы наблюдаем; объяснить же происхождение этого сходства мы сможем лишь тогда, когда нам удастся разделить генетическую и средовую общность родственников. Для этого в генетике человека применяют ряд экспериментальных схем, позволяющих разводить наследственное и средовое сходство и тем самым осуществлять количественную оценку наследуемости. Прежде чем подробно познакомиться с этими экспериментальными методами, рассмотрим, что лежит в основе генетического сходства между родственниками.
в начало
Общие гены у родственников. Понятие о вероятности. Коэффициент родства.
Мы знаем, что при оплодотворении и образовании зиготы происходит объединение хромосом и генов, находившихся в материнской и отцовской гаметах. В результате каждый ген индивида оказывается представленным двумя аллелями - одним материнским и одним отцовским. Таким образом, половина аллелей потомка получена им от матери и половина - от отца. Если в этой же семье рождается второй ребенок, то он также получает половину аллелей от матери и половину от отца, однако вследствие процессов рекомбинации, которые имеют место при образовании гамет, в зиготу попадает уже иной наследственный материал, другое сочетание аллелей, и второй ребенок практически никогда не обладает таким же генотипом, что и первый. Все же оба потомка одних и тех же родителей будут нести какую-то часть одинаковых генов. Число общих генов у потомков одних и тех же родителей определяется чистой случайностью, поскольку, как мы уже знаем, при образовании гамет в первом делении мейоза гомологичные хромосомы распределяются по гаметам случайным образом. Однако случайность не есть нечто неопределенное. Мерой случайности является вероятность.
Все мы сталкиваемся в жизни со случайными событиями. Подбрасывая монетку, мы знаем, что сторона, на которую она упадет, определяется случайностью, но если продолжить опыт с монеткой много раз, то мы заметим, что примерно в половине случаев выпадет "орел", а в половине - "решка". В случае с монеткой вероятность того, что выпадет орел, равняется одному шансу из двух возможных. Говорят, что в этом случае вероятность события составляет 1/2. Если бросать игральный кубик, то вероятность того, что выпадет одна из граней, например "шесть", составит 1/6. Вообще вероятность какого-либо события выражается в виде дроби, в числителе которой стоит число благоприятных шансов, а в знаменателе - общее число шансов всех исходов события. При образовании гамет также происходят вероятностные события. Когда мы рассматривали дигибридное скрещивание в опытах Г. Менделя, то видели, что с равной вероятностью могут образовываться четыре типа гамет. Иными словами, вероятность образования гамет с аллелями АВ, так же как и с остальными тремя сочетаниями аллелей (Ab, aB, ab), будет равна 1/4. В результате вероятностных событий, происходящих при образовании гамет, родные братья и сестры (сибсы) получают какое-то количество одинаковых аллелей. Поясним это на примере. Предположим, родители обладают различными аллелями одного и того же гена. Пусть у отца имеются аллели АВ, а у матери - СD. Гаметы отца могут нести либо аллель А, либо аллель В. Каждый из сибсов с равной вероятностью может получить как аллель А, так и аллель В. Понятно, что в среднем половина потомков будут нести аллель А и половина - аллель В. Соответственно вероятность того, что оба сибса будут обладать одним и тем же аллелем, составит 1/2. То же самое справедливо и для материнских аллелей - С и D (см. табл.):
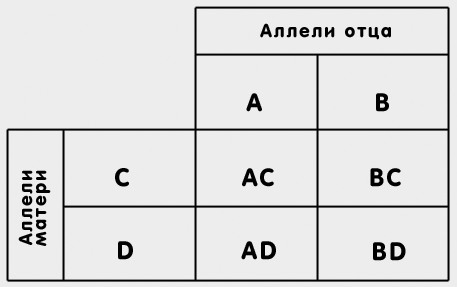
в начало
Сочетание материнских и отцовских аллелей у потомков
Вероятность того, что двое людей обладают одинаковыми аллелями, называется коэффициентом родства. Коэффициент родства соответствует доле идентичных аллелей, имеющихся у двух индивидов, благодаря их происхождению от общего предка. Коэффициенты родства рассчитываются теоретически на основе теории вероятностей и математической статистики. Приведенный пример является лишь наглядной иллюстрацией. Коэффициент родства для сибсов, как мы видели, составляет 1/2, т.е. в среднем у сибсов 1/2 генов идентичны и получены от одного общего предка. Такой же коэффициент родства характерен и для пар родитель-потомок. Приведенный пример может служить иллюстрацией и в этом случае. Понятно, что только половина потомков будут нести тот же аллель, что и у одного из родителей. В нашем примере коэффициент родства родителей равен 0, т.к. аллели матери и отца не совпадают. В реальной жизни, особенно при близкородственных браках (например, между двоюродными сибсами), родители также могут обладать одинаковыми аллелями. Так, коэффициент родства между двоюродными сибсами составляет 1/8. Это означает, что в среднем 1/8 генов получены ими от общего предка. Чем отдаленнее родство, тем меньше общих генов можно обнаружить в парах родственников. В таблице ниже приведены теоретически рассчитанные коэффициенты родства для различных пар родственников.

В большинстве культур браки между близкими родственниками запрещаются. Это связано с тем, что при близкородственных браках выше вероятность встречи рецессивных аллелей, связанных с различными аномалиями. В гомозиготном состоянии такие аллели приводят к возникновению патологических отклонений. При неродственных браках вероятность проявления патологических аллелей гораздо ниже.
в начало
Способы количественной оценки фенотипического сходства между родственниками
Итак, сходство между родственниками возникает, с одной стороны, за счет общих генов, а с другой стороны, за счет общей среды. Чтобы количественно оценить степень сходства в парах родственников, чаще всего используют конкордантность и корреляцию. В некоторых случаях пользуются также регрессией.
Конкордантность
При анализе сходства/различия альтернативных признаков используют оценки конкордантности. Чаще всего оценки конкордантности используются в клинической психогенетике при изучении причин различных психических заболеваний или отклонений в развитии. Пары родственников называются конкордантными, если оба имеют или не имеют данный признак. Соответственно, дискордантными называются пары, в которых один обладает данным признаком, а другой - нет. Для оценки конкордантности подсчитывается процент совпадения альтернативных признаков в парах родственников. Те индивиды, которые обладают интересующим нас признаком, называются пробандами.
Исследованию подлежат пробанды и их родственники различной степени родства (близнецы, сибсы, родители, дети и т.п.). Например, при изучении наследственности шизофрении в качестве пробандов могут подбираться больные близнецы, как монозиготные (МЗ), так и дизиготные (ДЗ). Предположим, в таком исследовании было получено, что для 20 пробандов (МЗ близнецов), больных шизофренией, в 15 случаях был болен и партнер пробанда, т.е. пары оказались конкордантными по шизофрении, тогда как в оставшихся 5 парах (дискордантных по шизофрении) партнеры оказались здоровыми. В данном случае конкордантность (С) равна: СМЗ = (15/20)100 = 75%.
В то же время из 20 пробандов ДЗ близнецов лишь у 10 оказался болен и другой член пары, т.е. на 10 конкордантных пришлось столько же дискордантных пар. В этом случае конкордантность ДЗ близнецов равна:
СДЗ = (10/20)100 = 50%.
В результате такого исследования можно сделать вывод, что риск заболевания шизофренией выше для родственников, связанных более тесным родством, т.е. имеющих больше общих генов. Если результаты подобных исследований оказываются статистически достоверными, можно предполагать наличие наследственной обусловленности заболевания.
Коэффициенты конкордантности помогают определить риск заболеваемости для различных категорий родственников. Например, если для сибсов конкордантность по шизофрении составляет 10%, можно считать, что риск заболевания шизофренией у человека, имеющего брата или сестру, страдающих этим заболеванием, составит 1 на 10 случаев. Как правило, конкордантность для родственников сравнивается с встречаемостью болезни в популяции. Например, если было обнаружено, что заболеваемость шизофренией для родственников первой степени родства составляет 10%, а в популяции встречаемость этой болезни всего 1%, можно говорить о более высоком риске заболевания у родственников. В таблице приведены частоты заболеваемости шизофренией для различных категорий родственников:
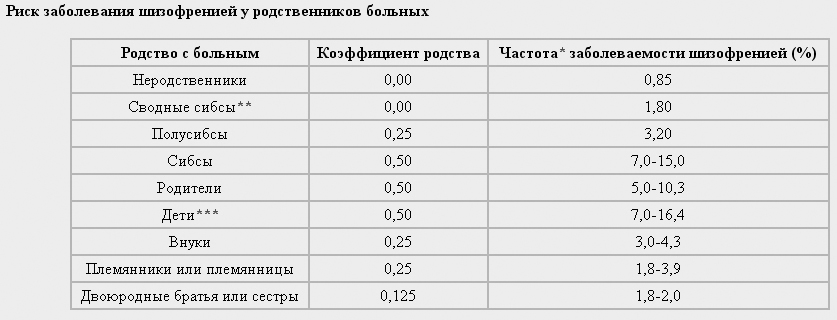
Сопоставление оценок конкордантности МЗ и ДЗ близнецов дает возможность оценить долю влияния наследственных факторов на вариативность исследуемого признака, т.е. коэффициент наследуемости h2. Для этого можно воспользоваться формулой Хольцингера:
h2=(СМЗ - СДЗ) / (100 – СДЗ) = (75 - 50) / (100 - 50) = 0,5
Итак, в приведенном примере коэффициент наследуемости (в широком смысле слова) равен 0,5.
Корреляция
При анализе количественных признаков сходство между родственниками оценивается с помощью корреляции. Мы уже упоминали об этом, когда обсуждали проблему ассортативности. Сейчас поговорим о корреляциях более подробно.
В статистике коэффициент корреляции обычно используется для оценки меры связи между двумя величинами. Так, можно, например, задать вопрос, существует ли связь между двумя количественными признаками у человека, например, между густотой волос и ростом. Здесь возможны три варианта:
1) чем гуще волосы, тем выше рост,
2) чем гуще волосы, тем рост ниже,
3) эти признаки никак не связаны между собой.
Если верно последнее утверждение, то мы бы сказали, что эти признаки не коррелируют. В первом случае можно говорить о положительной корреляции, во втором - об отрицательной. Основной принцип подсчета корреляции заключается в следующем. Возьмем, например, такие признаки, как рост человека и его вес. Предположим, что у группы индивидов мы провели соответствующие измерения и занесли данные в таблицу:
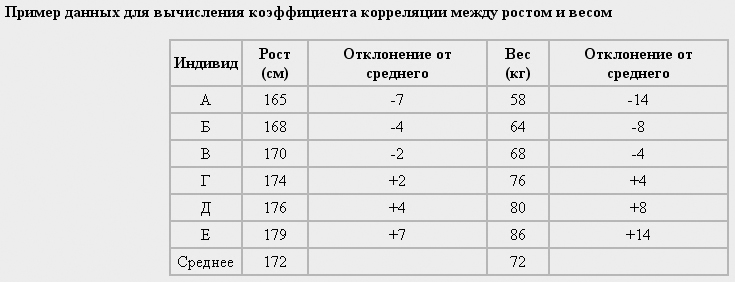
Рассматривая таблицу, мы можем заметить, что отклонение роста каждого человека от средней величины прямо пропорционально таким же отклонениям, относящимся к его весу. В этом примере рост и вес демонстрируют максимальную положительную корреляцию, величина которой равна +1,0. Если мы расположим значения в одном из столбцов в обратном порядке, то получим отрицательную корреляцию с величиной коэффициента, равной -1,0. Если мы в случайном порядке перемешаем все цифры, то корреляция будет близка к 0.
Графически положительная корреляция между двумя величинами может быть представлена в виде линии с положительным наклоном
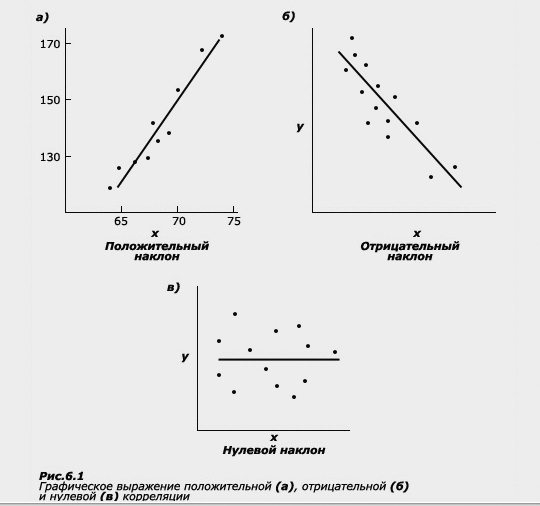
(рис. а), при этом на осях Х и Y откладываются значения коррелируемых признаков; отрицательная корреляция может быть представлена в виде линии с отрицательным наклоном (рис. б), отсутствие корреляции выражается в отсутствии наклона соответствующей линии (рис. в). Таким образом, величина корреляции говорит нам о том, насколько отклонения от средней одной величины совпадают с отклонениями другой. (Напомним, что все, что связано с отклонениями от средних величин - это область, близкая к проблемам вариативности и измерениям дисперсии.) Однонаправленный характер отклонений приводит к возникновению высокой положительной корреляции. Вместе с тем величина коэффициента корреляции не несет никакой информации об абсолютных величинах двух признаков. Взглянув на таблицу, мы убедимся, что в колонках цифр абсолютные значения роста и веса отличаются примерно на сто единиц. Две переменные могут идеально коррелировать друг с другом, даже если каждое значение одной значительно больше, чем каждое значение другой. Это обстоятельство имеет непосредственное отношение к пониманию значений корреляций в оценке сходства между родственниками.
При оценке сходства между родственниками измеряют не два признака у одних и тех же людей, а один и тот же признак в парах родственных индивидов. Ими могут быть близнецы, сибсы, родители и дети и даже неродственники, живущие в одной семье (имеются в виду семьи с приемными детьми) и т.д. Принцип же подсчета корреляций такой же. Важны не абсолютные величины признака, а отклонения от средней. Если отклонения однонаправленны, то и корреляция между родственниками будет высокой.
В зависимости от типа родственников используется тот или иной тип коэффициента корреляции. В тех случаях, когда оценивается сходство между парами родственников, принадлежащих разным поколениям (родитель-ребенок, дед-внук и т.д.), используют межклассовый коэффициент корреляции, предложенный Карлом Пирсоном.
Для оценки степени сходства между близнецами и сибсами используется внутриклассовый коэффициент корреляции

Итак, в генетике поведения мерой сходства между родственниками чаще всего является корреляция, которая не предполагает сходства в абсолютных величинах признака. Однако нередко в обыденном понимании сходство между родственниками отождествляется со сходством в абсолютных или средних величинах. Такое понимание сходства может приводить к неверной интерпретации получаемых результатов.
Рассмотрим гипотетический пример, который иллюстрирует возможность возникновения некоторых заблуждений по поводу роли наследственных и средовых факторов в возникновении индивидуальных различий.
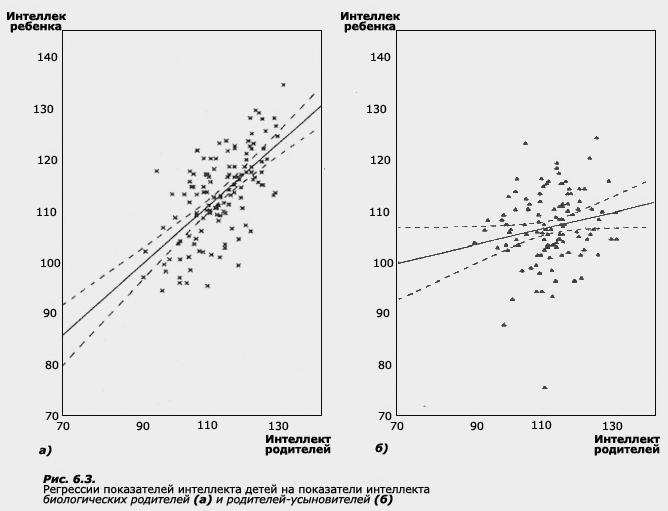
Предположим, что группа детей из бедных семей была усыновлена группой родителей из средних или состоятельных слоев общества. Приемные родители по своему интеллектуальному потенциалу и материальным возможностям смогли обеспечить детям идеальные возможности для развития. Допустим, что, когда дети выросли, было произведено измерение коэффициента интеллекта у самих детей, а также у их биологических и приемных матерей. Предположим, что данные этого измерения оказались следующими:

В настоящем примере получается идеально высокая положительная корреляция (равная +1) между детьми и их биологическими матерями, поскольку в обеих этих группах наблюдается однонаправленность отклонений от среднего, что и создает высокую положительную корреляцию. Корреляция с матерями-усыновителями будет гораздо ниже. Однако если мы взглянем на абсолютные величины IQ, то легко можем заметить, что приемные дети как группа гораздо ближе к группе их матерей-усыновителей. Действительно, средние величины интеллекта в этих группах совпадают, тогда как в группе биологических матерей они гораздо ниже. Поскольку приемные дети имеют общую среду с усыновившими их матерями и не имеют таковой с их биологическими матерями, а по абсолютным величинам IQ оказываются гораздо ближе к матерям-усыновителям, сам собой напрашивается вывод о средовой детерминации интеллекта. Вместе с тем высокая корреляция между биологическими родственниками и отсутствие таковой у людей, не связанных генетическим родством, свидетельствуют о высокой наследуемости признака. В этом примере содержится основной парадокс данных количественной генетики: высокая наследуемость вполне может сочетаться с чувствительностью конкретных генотипов к средовым изменениям. Усыновление в семьи с более благоприятной средой вполне может привести к тому, что дети будут опережать своих биологических матерей по уровню интеллекта. Вместе с тем легко заметить, что хотя дети оказались на другом уровне шкалы, они сохранили свои ранговые места в соответствии со своим биологическим происхождением. Таким образом, даже если индивидуальные особенности детей идеально коррелируют с особенностями их биологических родителей, эти дети как группа в среднем могут больше походить на родителей-усыновителей.
В нашем примере данные генетики поведения говорят о том, что вариативность интеллекта в популяции в основном обусловлена вариативностью генотипов, а не вариативностью сред. Это тот вывод, который может сделать генетик. Но психолог, измерив средние величины интеллекта в группах детей и их приемных и биологических родителей, имеет полное право сказать, что наблюдаемое сходство по интеллекту между приемными детьми и их родителями-усыновителями обусловлено общей средой (общая среда в нашем примере только у приемных детей и матерей-усыновителей). Как ни странно, но выводы генетика и психолога абсолютно не противоречат друг другу, но для генетика мерой сходства является корреляция, которая указывает на то, что показатели интеллекта у родственников в популяции ковариируют, а для психолога такой мерой является сходство в абсолютных значениях или средних величинах интеллекта в группах приемных детей и их матерей-усыновителей, которое говорит о возможности среды влиять на фенотип в индивидуальном развитии. Среда, влияя на индивидуальное развитие каждого конкретного ребенка, приводит к увеличению сходства в абсолютных величинах признака между детьми и родителями-усыновителями. Однако наблюдаемые межиндивидуальные различия по интеллекту в данном примере должны быть объяснены генетическими различиями между индивидами. Основанием для этого и является высокая корреляция между биологическими родственниками.
Регрессия
Коэффициент корреляции не предполагает наличия какой-либо причинно-следственной зависимости между переменными. Если с увеличением переменной х наблюдается рост переменной y, то мы констатируем наличие положительной корреляции между этими переменными, однако на этом основании мы еще не можем утверждать, что увеличение у является следствием увеличения х. Например, в городах мы можем обнаружить, что количество кафе и столовых положительно коррелирует с количеством больниц и поликлиник. Естественно, из этого не следует, что общественное питание является причиной высокой заболеваемости населения, что и приводит к появлению новых лечебных учреждений. Просто количество тех и других заведений определяется численностью населения города: чем больше жителей, тем больше необходимо как кафе, так и поликлиник. Отсюда и высокая положительная корреляция. Таким образом, коэффициент корреляции позволяет определить лишь наличие статистической связи между переменными, но не позволяет установить причину этой связи. В статистике существует и другой метод измерения связи, который предполагает оценку зависимости одной переменной от другой. Это метод линейной регрессии. Метод регрессии позволяет предсказать, какую величину будет иметь зависимая переменная у при любых значениях независимой переменной х. Речь фактически идет об уравнении регрессии
уi = a + b( xi - x ),
в котором нам необходимо определить величины а и b соответствующей линии регрессии. Независимая переменная (xi - x) представляет собой отклонение признака данного индивида от среднепопуляционной величины. Линия регрессии строится таким образом, чтобы квадраты расстояний между ней и всеми точками на графике были минимальными.
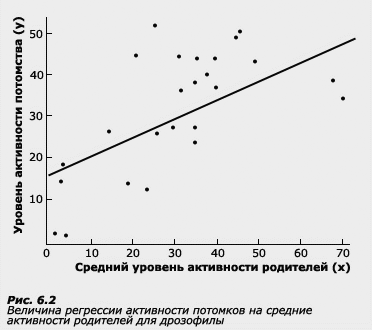
Коэффициент b называется коэффициентом регрессии у на х. Если коэффициент регрессии достоверно выше 0, то говорят о зависимости переменной у от переменной х. В количественной генетике регрессия применяется в основном в исследованиях родителей и детей. Часто используют одновременно и регрессию, и корреляцию. Регрессия имеет ряд преимуществ по сравнению с корреляцией, применение которой ограничено рядом условий, о которых речь пойдет в следующем разделе. Регрессия менее чувствительна к этим условиям.
Термин регрессия был введен Ф. Гальтоном при исследовании роста у родителей и детей. В этой работе Ф. Гальтон отметил, что у более высоких отцов сыновья также отличаются высоким ростом, но все же они несколько ниже своих отцов. У отцов небольшого роста сыновья так же невысоки, но они обычно выше своих отцов. Таким образом, рост детей как бы стремится к популяционной средней.
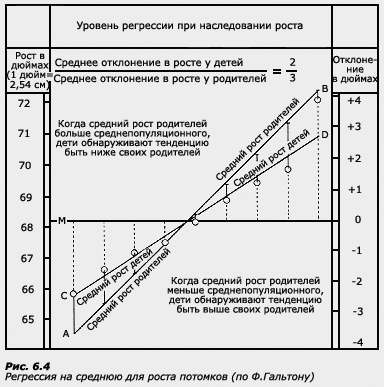
Это явление Ф. Гальтон назвал регрессией на среднюю.
в начало
Условия соответствия коэффициента корреляции коэффициенту родства
Как мы уже говорили, выявление степени внутрисемейного сходства является основным методом количественной генетики. Основоположники биометрической генетики, начиная с Ф. Гальтона, ввели этот метод в обиход генетических исследований, практически ничего не зная о механизмах, приводящих к появлению сходства между родственниками. Сейчас мы знаем, что в основе фенотипического сходства лежит сходство генетическое, обусловленное общностью генов, полученных от одного предка (если, конечно, признак не целиком обусловлен средой, а все же зависит от действия генов). В 1918 г. английский генетик и статистик Р. Фишер показал, что результаты, полученные основоположниками биометрической генетики, могут быть объяснены именно с позиций менделевской генетики. Можно сказать, что в этом году две независимо развивавшиеся ветви генетики - менделевская и гальтоновская - наконец нашли путь к объединению. Заслуга Р. Фишера в том, что он показал, что ожидаемая степень сходства между родственниками объясняется полигенным наследованием, т.е. детерминацией признака не одним, а многими генами, и при определенных допущениях, о которых речь пойдет ниже, она соответствует коэффициентам родства. В основе этого лежат вероятностные процессы, происходящие при расщеплении и рекомбинации аллелей, открытые Г. Менделем. Оказалось, что регрессия (напомним, что это термин Ф. Гальтона) родителя к ребенку или ребенка к родителю, или сибса к сибсу, или других пар родственников соответствует доле общих генов у этих родственников, т.е. коэффициенту родства. Коэффициент корреляции равен корню квадратному из произведения двух регрессий, т.е. регрессии первой величины по второй и второй величины по первой . Коэффициент корреляции будет эквивалентен регрессии, когда две регрессии (например, ребенка к родителю и родителя к ребенку) равны. Таким образом, при определенных условиях коэффициент корреляции в парах родственников должен соответствовать доле общих генов, или коэффициенту родства.
Каковы же эти условия, или допущения, при которых реально наблюдаемое внутрисемейное сходство должно соответствовать теоретически рассчитанному? Условия эти сводятся к следующему:
исследуемый признак детерминирован исключительно генотипом, условия среды никак не сказываются на фенотипе;
гены обладают чисто аддитивным эффектом, отношения доминантности и рецессивности, эффекты эпистаза не имеют места;
у мужа и жены отсутствует корреляция по изучаемому признаку, т.е. брак является случайным в отношении изучаемого признака. Иначе говоря, не наблюдается ассортативности по исследуемому признаку.
Итак, существует три условия, при точном соблюдении которых реально получаемые корреляции между родственниками должны совпадать с теоретически рассчитанными коэффициентами родства. Из всех количественных признаков человека лучше всего удовлетворяют этим условиям дерматоглифические узоры на пальцах. Каждый человек имеет свой индивидуально специфический характер дерматоглифических борозд.

Этот рисунок закладывается на ранних этапах эмбриогенеза и впоследствии не претерпевает никаких изменений, т.е. не испытывает средовых модификаций. Распределение индивидов по числу борозд соответствует нормальному, гауссову, следовательно, мы можем предположить, что здесь имеет место чисто аддитивное наследование. Браки по такому признаку, естественно, не могут быть избирательными.
В отличие от корреляции, регрессия менее чувствительна к ассортативности, поэтому регрессией можно пользоваться в семейных исследованиях, когда предполагается избирательность браков по изучаемому признаку. Регрессия позволяет также выявлять так называемый материнский эффект, т.е. влияние фенотипа матерей на фенотип потомства. При материнском эффекте регрессия потомков к матерям значительно больше, чем регрессия к отцам. По регрессии можно судить и о наличии эффектов доминирования. Обычно для изучения наследуемости стараются применять различные методы: вычисление коэффициентов корреляции между различными группами родственников (родители-потомки, сибсы, полусибсы, матери-дочери и т.д.) и вычисление коэффициентов регрессии тем или иным способом.
Выводы
Сходство между родственниками, проживающими вместе, возникает за счет общих генов и общей среды и, следовательно, включает в себя наследственный и средовой компоненты. Необходимо различать сходство семейное и сходство генетическое.
У родственников всегда имеются общие гены, полученные ими в силу происхождения от общих предков.
Число общих генов у потомков одних и тех же родителей определяется чистой случайностью. Мерой случайности является вероятность.
При образовании половых клеток (гамет) происходят вероятностные события. В результате родные братья и сестры получают какое-то количество одинаковых аллелей.
Вероятность того, что двое людей обладают одинаковыми аллелями, называется коэффициентом родства. Коэффициенты родства для различных категорий родственников рассчитываются теоретически на основе теории вероятностей и математической статистики.
В психогенетических исследованиях для количественной оценки сходства между родственниками пользуются коэффициентами конкордантности, корреляции и регрессии.
Коэффициент конкордантности используют при анализе сходства и различий между родственниками по альтернативным признакам, например по наличию или отсутствию какого-либо заболевания или отклонения. По коэффициентам конкордантности родственников разной степени родства можно судить о возможной наследуемости признака и риске заболевания для родственников.
Коэффициент корреляции используют при анализе сходства и различий между родственниками по количественным признакам. Высокая корреляция указывает на преобладание однонаправленных отклонений значения изучаемого признака у родственников от выборочной средней. Это не предполагает обязательного сходства в абсолютных величинах признака в парах родственников.
Коэффициент регрессии чаще всего применяется при исследованиях родителей и детей и, в отличие от коэффициента корреляции, может служить мерой причинно-следственной зависимости между переменными. Регрессия при определенных условиях соответствует доле общих генов у родственников, т.е. коэффициенту родства.
Если регрессии родителя к ребенку и ребенка к родителю совпадают, то коэффициент регрессии будет эквивалентен коэффициенту корреляции.
При определенных условиях теоретически рассчитанное сходство между родственниками (коэффициенты родства) совпадает с эмпирически полученными коэффициентами корреляции и регрессии. Эти условия таковы:
исследуемый признак является количественным и в его детерминации принимают участие только полигены, условия среды не влияют на признак;
гены обладают чисто аддитивным (суммирующимся) эффектом;
по данному признаку отсутствует ассортативность (избирательность браков).
В отличие от корреляции регрессия менее чувствительна к ассортативности.
в начало
Экспериментальные схемы генетико-популяционных исследований
Методы, базирующиеся на принципах генетической эпидемиологии, помогают генетикам разделить фенотипическую вариативность изучаемых признаков на генетические и средовые компоненты. В теме 5 мы подробно анализировали компоненты, на которые может быть разложена фенотипическая дисперсия в популяции. Наиболее простым и часто единственным решением является выделение суммарного компонента генетической изменчивости, представленного коэффициентом наследуемости (h2) и двух средовых составляющих - общей (с2) и различающейся (е2) среды. Для количественной оценки указанных компонентов исследователи чаще всего обращаются к двум экспериментальным схемам: исследованиям близнецов и семей с приемными детьми. Оба методических варианта могут использоваться в комплексе друг с другом и с семейным методом. Последний из методов - семейный - исторически возник раньше, чем два других. Его применил Ф. Гальтон, исследуя наследственность таланта. Однако семейный метод обладает слабой разрешающей возможностью для разделения наследственных и средовых компонентов дисперсии, поскольку наблюдаемое сходство между членами семьи может равным образом объясняться общностью генов и общностью среды, если родственники проживают совместно. Рассмотрим эти методы более подробно.
в начало
Близнецы и близнецовый метод
Близнецовый метод является наиболее распространенным в генетике поведения человека. Чтобы лучше представлять себе основы этого метода, сначала поговорим о явлении близнецовости вообще.
Биология близнецовости
Большинство млекопитающих рождают больше одного детеныша в помете. Это происходит потому, что во время овуляции у этих животных созревают и оплодотворяются сразу несколько яйцеклеток. У некоторых видов животных (лошади, крупный рогатый скот, высшие обезьяны) и человека при овуляции обычно образуется лишь одна яйцеклетка, и в результате рождается только один детеныш. Правда, иногда бывают исключения - одновременно созревают и оплодотворяются две, а иногда три и более яйцеклеток. В этом случае рождаются два или более детенышей, а поскольку они происходят из разных оплодотворенных яйцеклеток, или зигот, их называют дизиготными (ДЗ) близнецами, или двойняшками. Если рождается тройня, то правильнее назвать таких детей тризиготными близнецами. ДЗ близнецы не обязательно должны иметь одного отца. Если в период овуляции женщина имела контакт с разными мужчинами, вполне вероятно, что родившиеся ДЗ близнецы будут иметь разных отцов. Такие случаи описаны, в том числе и такие, когда один из младенцев был черным, а другой - белым.
Не все многоплодные беременности приводят к рождению ДЗ близнецов. В ряде случаев такие беременности приводят к рождению другого типа близнецов. Это монозиготные (МЗ) близнецы. МЗ близнецы принципиально отличаются от ДЗ, т.к. происходят не из разных зигот, а из одной и той же, которая на определенной стадии дробления по непонятным причинам разделилась на два самостоятельных организма. Происхождение МЗ близнецов из одной зиготы предопределяет абсолютную идентичность их генетической конституции. МЗ близнецы - это единственные люди на Земле, обладающие одинаковыми наборами генов. Вот почему они обычно как две капли воды бывают похожи друг на друга. Если разделение эмбриона на два организма произошло не полностью, могут родиться сросшиеся, или сиамские, близнецы. Часто такие близнецы погибают сразу после рождения, но в некоторых случаях выживают и даже могут быть отделены один от другого путем оперативного вмешательства. Название "сиамские" такие близнецы получили в честь первой изученной и описанной пары сросшихся близнецов. Это были сросшиеся близнецы Чанг и Энг, родившиеся в Сиаме (Таиланд) в 1811 г. Основную часть своей сознательной жизни они провели в Соединенных Штатах Америки. Оба были женаты и имели детей, один - 12, а другой - 10. Братья прожили 63 года, причем причиной их почти одновременной смерти послужила болезнь одного из братьев.

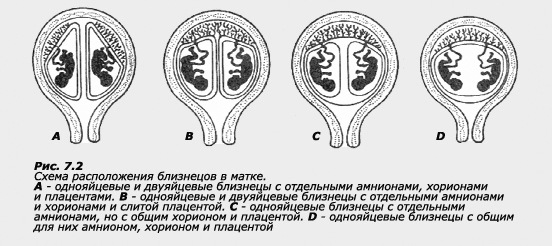
При рождении близнецов можно обнаружить четыре различных варианта в соотношении плодных оболочек:
1) близнецы имеют раздельные амнионы, хорионы и плаценты;
2) близнецы имеют раздельные амнионы и хорионы, но общую плаценту;
3) близнецы имеют раздельные амнионы, но общие хорион и плаценту;
4) близнецы имеют общие амнион, хорион и плаценту.
Различают три основных вида оболочек плода: амнион, хорион и плаценту. Амнион и хорион являются зародышевыми оболочками, т.е. развиваются из клеток зародыша. Амнион представляет собой внутреннюю зародышевую оболочку. Это тонкая оболочка, которая несет защитные функции и выделяет амниотическую жидкость. По мере роста зародыша амнион расширяется и всегда бывает прижат к стенке матки. Наружная зародышевая оболочка называется хорионом. Хорион играет важную роль в питании зародыша. Ворсинки хориона образуют тесную связь с материнским организмом через плаценту. Плацента - это временный орган, образующийся у плацентарных млекопитающих и происходящий одновременно из клеток плода и матери. Плацента служит связующим звеном между кровеносными системами матери и плода. После родов плацента, соединенная с хорионом (детское место), отделяется от стенки матки и также выходит через влагалище.
Для ДЗ близнецов характерны только два первых типа, тогда как при рождении МЗ близнецов могут наблюдаться все четыре типа соотношения плодных оболочек. Часто матери судят о зиготности своих детей-близнецов по количеству детских мест во время родов, но, как мы видим, такие суждения могут быть и ошибочными, поскольку здесь возможны различные варианты.
Частота рождения близнецов в разных популяциях различна, но эта разница возникает в основном за счет различий в частоте рождения ДЗ близнецов, тогда как количество рождений МЗ близнецов во всех популяциях примерно постоянно. Количество родившихся МЗ и ДЗ близнецов определяют приблизительно, используя очень простой способ подсчета. МЗ близнецы всегда однополые, а среди ДЗ - однополые и разнополые рождаются примерно с одинаковой вероятностью, следовательно, частота рождения ДЗ близнецов будет равна удвоенной частоте рождения разнополых двоен. А частота рождения МЗ близнецов, таким образом, составит разницу между частотой рождения всех близнецов вообще и частотой рождения ДЗ близнецов.
Допустим, общее число близнецовых родов за какой-то период составило N, из них разнополых двоен оказалось - n. Поскольку разнополые двойни - это всегда ДЗ близнецы, а рождение разнополых и однополых ДЗ близнецов происходит примерно с одинаковой частотой, тогда общее число ДЗ близнецов составит 2n. Понятно, что оставшаяся часть будет приходиться на долю МЗ близнецов, и их число будет определяться как разность между N и 2n, т.е. число МЗ близнецов составит (N-2n).
Хотя эти подсчеты дают лишь приблизительное представление об относительном количестве МЗ и ДЗ близнецов, все же таким способом проводят статистическую оценку рождаемости МЗ и ДЗ близнецов в разных странах. Оказалось, например, что в Соединенных Штатах Америки больше всего ДЗ близнецов рождается среди негритянского населения (110 пар на 10 000 родов), тогда как у американских китайцев и японцев они рождаются намного реже (порядка 20 на 10 000 родов). Вообще для монголоидных популяций характерна самая низкая частота многоплодия. Однако частота рождения МЗ близнецов у них несколько выше, чем в основной массе популяций. Например, у американских китайцев она достигает 48 на 10 000 родов, тогда как в большинстве стран колеблется в пределах 30-40. В Европе частота дизиготности составляет от 60 до 90 на 10 000 родов, а монозиготности - колеблется в пределах 30-40.
В нашей стране близнецы рождаются примерно с той же частотой, что и в Европе. ДЗ близнецов рождается в два раза больше, чем МЗ. Общее количество близнецов во всем мире составляет около 60 миллионов.
Вероятность рождения близнецов тем выше, чем больше номер беременности (т.е. какая беременность по счету) - близнецы чаще рождаются в поздних беременностях. Это правило касается исключительно ДЗ близнецов. Влияние возраста матери объясняется, по-видимому, повышением с возрастом уровня гонадотропина у женщин. Это и приводит к более частой полиовуляции. Эту гипотезу подтверждают и факты повышенной частоты многоплодных родов у женщин, проходивших лечение от бесплодия с помощью гонадотропных гормонов. В отношении дизиготного многоплодия имеются также факты, свидетельствующие о влиянии генетических факторов на вероятность рождения близнецов. Вероятность рождения ДЗ близнецов выше для тех женщин, родственники которых уже имели близнецов. Возможно, основной генетически детерминированной причиной в этом случае также может быть уровень гонадотропина. В отношении МЗ близнецов таких данных не имеется.
Феномен близнецовости во все времена привлекал внимание людей. О близнецах сложены легенды и сказания, близнецы являются героями многих произведений художественной литературы. Существует и специальная наука, изучающая феномен близнецовости. Она носит название гемеллология, что по-русски можно перевести как близнецеведение. За рубежом постоянно издается научная и научно-популярная литература о близнецах и для близнецов. К сожалению, на русском языке такой литературы немного. Многие издания давно стали библиографической редкостью. Для пользователей сети интернет недавно созданы отечественные сайты, в которых можно найти информацию о близнецах: twinsclub.ru и twins.org.ru
Концепция близнецового метода
Поскольку МЗ близнецы развиваются из одной зиготы и, следовательно, генетически идентичны, все наблюдаемые различия в их фенотипах должны объясняться исключительно средовыми причинами. Поэтому, чтобы определить, в какой мере изменчивость признака детерминирована генетическими факторами, казалось бы, достаточно измерить уровень сходства МЗ близнецов. И это действительно было бы так, если бы с момента рождения МЗ близнецы были разлучены и воспитывались в разных средах. Однако в большинстве случаев такого не происходит: близнецы воспитываются в одной и той же среде, поэтому, чтобы каким-то образом учесть влияния общей среды, в качестве контроля используют пары ДЗ близнецов, на которых фактор общей среды действует в той же мере, как и на МЗ близнецов, но зато общих генов у ДЗ близнецов примерно в два раза меньше, ведь по своей генетической конституции они соответствуют обычным братьям и сестрам. Соответственно уровень генетически обусловленного сходства ДЗ близнецов должен быть ниже, как у более отдаленных родственников, чем МЗ.
Идея близнецового метода была сформулирована Ф. Гальтоном еще в 1865 г., но окончательная его модификация связана с именем Г. Сименса. Именно Г. Сименс в 1924 г. заложил основы использования близнецового метода в генетике. Во-первых, он разработал надежный метод диагностики зиготности, ведь на глаз не всегда легко определить, являются ли близнецы МЗ или ДЗ парой. Метод диагностики зиготности Г. Сименса носит название метода полисимптомного сравнения и предполагает оценку сходства и различия близнецов по множеству параметров. Каждый из этих параметров в отдельности может ничего не говорить о зиготности, но измерение их в совокупности позволяет проводить более надежную диагностику. Во-вторых, Г. Сименс предложил исследовать не только МЗ близнецов, но и использовать в качестве контроля ДЗ пары. Практически после работ Г. Сименса принципы близнецового метода не претерпели значительных изменений.
Близнецовый метод в его классическом варианте основан на ряде допущений. Первое и главное из них - это равенство сред для партнеров как в парах МЗ, так и в парах ДЗ близнецов. Если изменчивость признака целиком определяется средой, то и МЗ, и ДЗ близнецы должны иметь по этому признаку одинаково высокие внутрипарные корреляции, по величине приближающиеся к 1,0. Если же изменчивость признака целиком зависит от генотипа, то коэффициент корреляции в группе МЗ близнецов окажется близким к 1,0, а корреляция в группе ДЗ, очевидно, будет приближаться к 0,5 (это коэффициент родства ДЗ близнецов), поскольку у ДЗ в среднем половина общих генов. Так, в идеале, работает близнецовый метод:

На основе этой схемы легко понять, как примерно оцениваются показатели h2, c2 и е2 (наследуемости, общей и различающейся среды). В предыдущем разделе приводилась формула Игнатьева для оценки показателя наследуемости: h2 = 2 (RМЗ - RДЗ).
Действительно, если подставить значения коэффициентов корреляции, то максимальный показатель наследуемости, равный 1, мы получим в том случае, когда вариативность признака целиком определяется генами. В реальных ситуациях таких высоких коэффициентов практически не бывает, поскольку вариативность признака определяется не только генами, но и средой, причем различающаяся среда, как мы уже знаем, дает весьма существенный вклад в изменчивость психологических характеристик.
Р. Пломин предложил на основе коэффициентов корреляции между близнецами оценивать также и вклад параметров общей и различающейся среды. Будем рассуждать так. Коэффициент корреляции МЗ близнецов отражает их генетическую и средовую общность. Поэтому мы можем его представить в виде суммы:
RМЗ = h2 + c2.
Отсюда находим с2: c2 = RМЗ - h2.
Чтобы найти е2, примем всю фенотипическую вариативность за 1. Она складывается из генетической (h2), общей средовой (с2) и различающейся средовой (е2), т.е. 1 = h2 + c2 + е2.
Далее остается на основе ранее вычисленных значений h2 и c2 получить е2 : е2 = 1 - (h2 + c2).
Запишем теперь все формулы окончательно:
h2 = 2(RМЗ - RДЗ) с2 = RМЗ - h2 е2 = 1 - (h2 + c2).
Таким образом, располагая коэффициентами корреляции, полученными в группах МЗ и ДЗ близнецов, мы можем примерно оценить вклад основных компонентов в фенотипическую дисперсию признака, не прибегая к сложным математическим процедурам, требующим специальных компьютерных программ. Иногда исследователи ограничиваются этими простыми вычислениями, но, как правило, только на первом этапе работы.
Второе основное допущение близнецового метода - это отсутствие систематических различий между близнецами и одиночнорожденными, иначе выводы, полученные в близнецовом исследовании, нельзя переносить на всю популяцию. Точно так же не должно быть систематических различий между самими типами близнецов - МЗ и ДЗ.
Для того чтобы понять, каким образом оценивается роль генотипа и среды по данным, получаемым близнецовым методом, каковы его возможности и ограничения, рассмотрим рисунок,
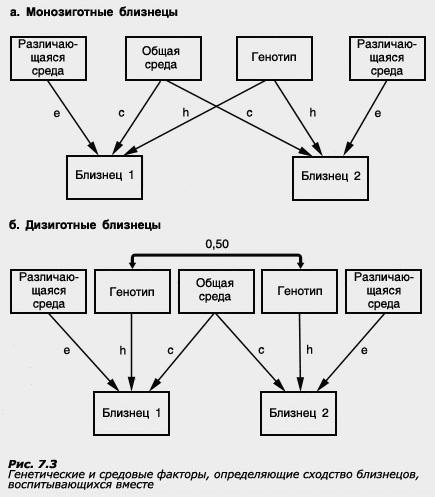
на котором представлен простейший вариант модели путей, т.е. способа, позволяющего схематически изобразить, каким образом генетические и средовые влияния определяют особенности фенотипа.
В верхней части рисунка (а) и (б) представлены те параметры, которые не регистрируются в близнецовом эксперименте. К ним относятся генотип, общая среда (та среда, которая совпадает у членов пары, например, экономический уровень семьи, психологические особенности родителей, общие друзья) и различающаяся среда (включающая в себя ту среду, которая не совпадает у членов пары, например, разные увлечения, разное отношение родителей, а также ковариация генотипа и среды и ошибка измерения). Генотип, общая и различающаяся среда представляют собой латентные переменные: мы только предполагаем их существование, но не измеряем их непосредственно в процессе экспериментального исследования.
В нижней части рисунков (а) и (б) (близнец 1 и близнец 2) показаны параметры, которые регистрируются в исследовании, - например, показатели интеллекта первого и второго близнеца.
Стрелками показаны пути от латентных переменных к изучаемому психологическому параметру - те влияния, которые оказывают генотип, общая и различающаяся среда на изучаемую характеристику. Путь от генотипа обозначен буквой "h" (от слова heredity - наследственность), путь от общей среды - буквой "с" (от слова common - общая), и путь от различающейся среды - буквой "е" (от слова error - ошибка; название связано с тем, что оценка различающейся среды включает в себя не только дисперсию средовых эффектов, не совпадающих у членов пары, но и ошибку измерения, т.е. неизбежные погрешности, которые бывают в эксперименте и которые тоже не совпадают для членов пары).
Как видно по рисунку, корреляции между членами МЗ и ДЗ пар могут определяться генотипом и общей средой. Если общая среда одинакова для партнеров МЗ и ДЗ пар близнецов, то сравнение внутрипарного сходства МЗ и ДЗ близнецов позволит получить вполне определенную информацию о роли генотипа и среды в вариативности изучаемой характеристики. Если постулат о равенстве средовых условий развития МЗ и ДЗ близнецов нарушается, то оценки компонент фенотипической дисперсии (наследуемость, дисперсии эффектов общей и различающейся среды) искажаются. Рассмотрим, в каких случаях такие искажения могут происходить.
Во-первых, средовые условия могут увеличивать внутрипарное сходство МЗ близнецов. В семье часто создают условия, усугубляющие их сходство (одинаково одевают, дают похожие имена и так далее). Подчеркивание сходства близнецов окружающими может приводить к возникновению дополнительного негенетического сходства между партнерами, что противоречит допущению о равенстве сред в МЗ и ДЗ парах, поскольку для ДЗ близнецов такое подчеркивание сходства менее характерно. Если мы изучаем признак, мало чувствительный к специфическим особенностям близнецовой среды (например, психофизиологические характеристики), то такие погрешности могут не оказать заметного влияния на сходство и различия близнецов. Если же признак чувствителен к такого рода средовым влияниям, то близнецовый метод оказывается мало пригоден для его изучения, т.к. основное допущение о равенстве сред оказывается неверным. Показатель наследуемости, вычисляемый для признака, чувствительного к особенностям близнецовой среды, будет завышенным, так как общая среда будет вносить больший вклад в сходство МЗ близнецов, чем в сходство ДЗ близнецов.
Во-вторых, средовые условия могут уменьшать внутрипарное сходство ДЗ близнецов. В целом ряде исследований показано, что средовые условия развития имеют тенденцию увеличивать различия ДЗ близнецов: родители склонны акцентировать несходство ДЗ близнецов (например, успехи в разных сферах деятельности); сами ДЗ близнецы стремятся подчеркнуть свою непохожесть (выбирают различные сферы деятельности, предпочитают иметь разных друзей). Это приводит к эффекту диссимиляции - постепенному увеличению различий ДЗ близнецов. Если изучаемая психологическая характеристика формируется при участии способствующих диссимиляции средовых особенностей, то показатель наследуемости будет так же завышен, как и в первом случае, поскольку общая среда будет вносить меньший вклад в сходство ДЗ близнецов, чем в сходство МЗ близнецов.
В-третьих, условия развития могут равным образом уменьшать сходство партнеров и МЗ, и ДЗ пар. Часть их связана с периодами внутриутробного развития и родов, часть - приходится на постнатальное развитие близнецов.
Во время внутриутробного развития очень часто близнецовые партнеры оказываются в неравноценных условиях. Все питательные вещества и кислород развивающиеся плоды получают через плаценту. Все ДЗ близнецы и примерно 1/3 МЗ близнецов имеют раздельные хорионы и плаценты. Оставшиеся 2/3 МЗ близнецов имеют общие хорион и плаценту. Их называют монохорионными близнецами (в отличие от МЗ близнецов, имеющих две плаценты и два хориона, которых называют дихорионными). В плодных оболочках монохорионных близнецов образуются различные соединения между сосудистыми системами одного и второго близнеца. Их называют шунтами. Наиболее опасны так называемые артерио-венозные шунты, т.е. соединения артерии одного близнеца с веной другого. При этом одному из близнецов может недоставать артериальной крови, богатой кислородом и питательными веществами, а у другого может быть ее избыток, что также не способствует нормальному развитию. Чаще всего возникает несколько примерно равных по мощности шунтов, которые компенсируют друг друга, но в некоторых случаях компенсация бывает недостаточной, и это приводит к тому, что один из близнецов развивается в условиях постоянного дефицита необходимых веществ. В результате такие близнецы сильно отличаются один от другого при рождении, в первую очередь по весу. Разница в весе при рождении может наблюдаться также у ДЗ близнецов и дихорионных МЗ. Такая разница может быть связана с неравномерным сдавливанием плацент при многоплодной беременности.
Второй этап развития, на котором также возникают обусловленные средой различия между близнецами - это процесс родов. Близнец, рождающийся первым, проходит через неподготовленные родовые пути и имеет больший шанс получить травму, но второй близнец сталкивается с целым рядом других трудностей. Он чаще занимает в матке неправильное положение (поперечное или тазовое предлежание), и, чтобы извлечь его, врачам чаще приходится применять искусственное родовспоможение. Кроме того, второй близнец дольше находится в родах и чаще испытывает кислородное голодание, что неблагоприятно сказывается в первую очередь на развитии центральной нервной системы. По данным статистики, смертность "вторых" близнецов больше, чем смертность "первых". Таким образом, различия в качестве кровоснабжения во внутриутробном периоде и неравноценность условий при прохождении родовых путей могут привести к существенным средовым различиям между близнецами сразу же после рождения.
Средовые различия между близнецами возникают и в постнатальном периоде, даже при условии воспитания в одной семье, т.е., казалось бы, при полной общности среды. К возникновению таких различий чаще всего приводит предвзятое отношение родителей к каждому из близнецов. Если дети родились с разницей в весе, если один из близнецов более ослаблен и т.п., то между детьми сразу же наблюдаются врожденные отличия. Далее все зависит от отношения родителей. Если родители придают этим различиям особое значение и начинают по-разному обращаться с каждым из близнецов (часто более слабого ограждают от физических нагрузок, больше балуют, меньше наказывают и т.д.), то врожденные различия в физическом развитии дополнятся и различиями в развитии психики. Нередко при врожденных различиях между близнецами родители как бы делят детей между собой - более слабого опекает мать, а с более сильным больше занимается отец. Все эти особенности воспитания только усугубляют уже существующие различия и, по большей части, не способствуют правильному развитию детей. Еще один фактор, часто приводящий к возникновению психологических различий между близнецами в семье, - это разделение ролей между близнецами. Как и в любой группе, в паре может выделиться руководитель (лидер), второй близнец будет занимать подчиненное положение. Между партнерами могут образоваться и комплементарные отношения. Известны случаи, когда один из близнецов всегда выполняет одни обязанности, а второй - другие. Например, в одной из подобных пар одна сестра, делая уроки, всегда читала текст вслух, а другая только слушала. В результате, уже будучи взрослой, одна из девушек испытывала трудности в чтении.
Таким образом, если средовые условия оказывают различное влияние на формирование изучаемой характеристики у МЗ и ДЗ близнецов, то показатель наследуемости этой характеристики может оказаться искаженным: заниженным, если общая среда вносит меньший вклад в сходство МЗ близнецов, чем в сходство ДЗ близнецов; завышенным - в противоположном случае.
Разновидности близнецового метода
Существует несколько вариантов работы с близнецами, которые используются для различных целей.
1) Классический близнецовый метод. К классическому варианту близнецового метода относится такая схема, при которой выраженность исследуемого признака сопоставляется в парах МЗ и ДЗ близнецов и оценивается уровень внутрипарного сходства партнеров. Принцип применения этого варианта близнецового метода был разобран в предыдущем разделе.
2) Метод контрольного близнеца. Используется на выборках МЗ близнецов. Поскольку МЗ близнецы очень сходны по многим признакам, из них можно составить две выборки, уравненные по большому числу параметров. Такие выборки используют для изучения влияния конкретных средовых воздействий на изменчивость признака. Для этого отбирается по одному МЗ близнецу из каждой пары для специфического воздействия (двигательная тренировка, экспериментальное обучение, лечебные процедуры и т.п.). Остальные близнецы такому воздействию не подвергаются и служат контрольной группой. Таким способом можно, например, сравнивать эффективность разных программ обучения. Поскольку в эксперименте принимают участие генетически идентичные люди, можно считать такой способ сравнения моделью для изучения воздействия разных факторов среды на одного и того же человека.
3) Лонгитюдное близнецовое исследование. При этом варианте близнецового метода проводится длительное наблюдение - прослеживание одних и тех же близнецовых пар. Фактически это сочетание классического близнецового метода с лонгитюдным. Метод открывает широкие возможности для изучения генетических и средовых факторов в развитии. В настоящее время широко применяется и в зарубежных, и в отечественных исследованиях.
4) Метод близнецовых семей. Представляет собой сочетание близнецового метода с семейным. При этом исследуются члены семей взрослых близнецовых пар. Дети МЗ близнецов по своей генетической конституции являются полусибсами, т.е. как бы детьми одного и того же человека от разных браков. Этим методом, изучая мужские и женские пары МЗ и их потомство, можно исследовать, например, влияние материнского эффекта. Этот метод широко используют также при изучении наследственных причин ряда заболеваний, при которых один из МЗ близнецов может быть поражен болезнью, а другой - нет. Например, было показано, что при шизофрении эмпирический риск заболевания примерно одинаков для родственников МЗ близнецов, независимо от того, поражены ли оба партнера или только один из них.
5) Метод разлученных близнецов. Из-за особенностей развития МЗ и ДЗ близнецов классический близнецовый метод и его варианты принято считать "нежесткими" экспериментальными схемами: в них невозможно абсолютно однозначно развести влияние генетических и средовых факторов, поскольку условия развития МЗ и ДЗ близнецов по целому ряду особенностей оказываются несопоставимыми.
Вследствие этого результаты исследований, проводящиеся по "нежестким" схемам, требуют дополнительной валидизации. Она может быть двоякого рода. Во-первых, в процессе проведения исследования можно проверять правильность постулата о сходстве среды МЗ и ДЗ близнецов, т.е. доказывать, что на данную изучаемую характеристику не влияют различия в условиях развития МЗ и ДЗ близнецов. Сделать это довольно трудно, т.к. не всегда можно точно определить, что влияет на формирование той или иной психологической характеристики, а что - нет, но учет по крайней мере известных влияний повышает надежность результатов. Во-вторых, результаты таких исследований можно сопоставлять с результатами, полученными в экспериментах, проведенных по "жестким" схемам, которые позволяют надежно развести влияние средовых и генетических факторов. К числу таких экспериментальных схем относится метод разлученных близнецов.
Метод разлученных близнецов предполагает сравнение внутрипарного сходства близнецов, которые были разлучены в раннем возрасте и никогда не встречались друг с другом. Если МЗ близнецы, имеющие абсолютное генетическое сходство, растут в разных условиях, ходят в разные школы и не имеют никаких возможностей оказывать друг на друга влияние, то их сходство будет полностью результатом их генетической идентичности, а несходство - целиком определяться их средовыми условиями. Однако считать разлученных близнецов полностью свободными от влияний общей среды неправомерно, поскольку они всегда разделяют общие условия внутриутробного развития, которые могут привести к негенетическому внутрипарному сходству. Несмотря на все преимущества, этот вариант близнецового метода также не является идеальной жесткой схемой.
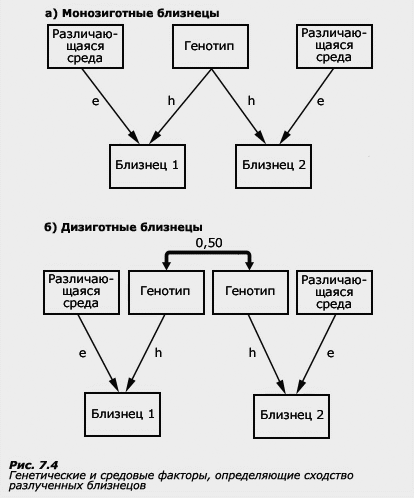
Все же эта схема близнецового метода (рис. а и б) дает возможность сделать более надежные выводы о генетических и средовых источниках индивидуальных различий, но по причинам, о которых нетрудно догадаться, ее крайне трудно реализовать. Разделение детей, родившихся в одной семье, обычно является следствием особых, и как правило, трагических обстоятельств - смерти родителей или их полной экономической несостоятельности, не позволяющей прокормить членов своей семьи. К счастью, такие случаи достаточно редки, но соответственно невелики и возможности для применения метода разлученных близнецов: даже в тех исследованиях, авторы которых собирали разлученных близнецов по всему миру, насчитывается немногим более 100 пар.
В выборках разлученных близнецов, описанных к настоящему времени, в очень широких пределах варьируют возраст испытуемых (от подросткового до преклонного); возраст, в котором близнецы были разлучены; время, которое прошло с тех пор, как близнецы нашли друг друга. Кроме того, разлучение часто оказывается неполным, а средовые условия, в которых живут близнецы, - похожими (например, близнецы живут у разных родственников в сходных условиях и время от времени встречаются). Все это вносит в исследования разлученных близнецов разнообразные искажения, как правило увеличивающие негенетическое сходство разлученных МЗ близнецов и снижающие разрешающие возможности этого метода. Добавим также, что негенетическое сходство может увеличиваться и за счет общего материнского эффекта в период внутриутробного развития, когда многие факторы среды, например характер питания матери, ее эмоциональное состояние, характер протекания беременности и т.д., являются общими для обоих близнецов.
6) Метод частично разлученных близнецов. В последнее время в генетических исследованиях стал применяться метод, который состоит в сравнении внутрипарного сходства МЗ и ДЗ близнецов, живущих какое-то время врозь.
Став взрослыми, близнецы вступают в брак, разъезжаются по разным городам. Их контакты становятся эпизодическими. Если близнецы, живущие долгое время врозь, не отличаются по внутрипарному сходству от близнецов, живущих вместе, можно сделать вывод, что средовые условия не изменяют степени сходства близнецов. Если сходство близнецов друг с другом по какой-либо психологической характеристике уменьшается в зависимости от продолжительности того периода, в течение которого близнецы живут врозь, можно сказать, что средовые условия воздействуют на изучаемую характеристику.
В исследовании частично разлученных близнецов можно также определить, в какой мере соблюдается постулат о равенстве сред МЗ и ДЗ близнецов. Так, если МЗ близнецы, живущие врозь, становятся все менее похожи друг на друга по какой-то психологической характеристике, а ДЗ близнецы, живущие врозь, не отличаются по внутрипарному сходству от ДЗ близнецов, живущих вместе, можно сделать вывод, что средовые условия МЗ и ДЗ близнецов неравноценны, а выводы о наследуемости изучаемой характеристики, полученные в классических близнецовых исследованиях, завышают показатель наследуемости этой характеристики.
Далее мы приводим еще три экспериментальные схемы, которые, во-первых, важны для изучения явления близнецовости самого по себе (напомним, что наука о близнецах носит название гемеллологии), а во-вторых, имеют значение как обслуживающие близнецовый метод, поскольку некоторые особенности развития и взаимоотношений близнецов могут сказываться на оценках наследуемости.
1) Исследование близнецов как пары - предполагает исследование специфических близнецовых эффектов и особенностей внутрипарных отношений. Используется как вспомогательный метод для проверки справедливости допущения о равенстве средовых условий для партнеров МЗ и ДЗ пар.
2) Исследование одиночных близнецов. Предполагает сопоставление особенностей развития одиночнорожденных детей и детей, родившихся в многоплодных родах, при которых один из партнеров умер при рождении. Также является вспомогательным методом, позволяющим оценить влияние пренатальных особенностей развития близнецов на их дальнейшее развитие.
3) Сопоставление близнецов с неблизнецами. Также вспомогательный метод, позволяющий оценить существенность различий между близнецами и неблизнецами. Если близнецы и все остальные люди значимо не различаются, т.е. принадлежат к одной генеральной совокупности, данные близнецовых исследований могут быть распространены на всю популяцию. Несколько поясним назначение этого метода.
Цель близнецовых исследований состоит в получении данных, применимых не только к самой выборке близнецов, но и ко всей популяции в целом. Чтобы выводы, сделанные в близнецовом исследовании, были справедливы и по отношению к одиночнорожденным, необходимо учитывать систематические различия между близнецами и неблизнецами.
Основная масса различий между близнецами и одиночнорожденными детьми связана с периодами раннего развития. Близнецы чаще рождаются недоношенными, у них чаще встречаются врожденные аномалии развития, их вес при рождении в среднем ниже веса одиночнорожденных детей, что отчасти объясняется большей частотой недоношенности. Частота мертворождений и детской смертности для близнецов также выше. У близнецов выше риск умственной отсталости. В среднем интеллект близнецов младшего возраста несколько ниже, чем у их сверстников-одиночнорожденных. По данным известного исследователя близнецов и близнецовости Рене Заззо, среднее значение коэффициента интеллекта (IQ) у близнецов ниже среднепопуляционного на 7 баллов. Чаще всего у близнецов оказываются сниженными вербальные способности.
Когда проводили сопоставление выборки близнецов, чьи партнеры умерли в раннем детстве (т.е. практически близнецы из этой выборки воспитывались как одиночки), с выборкой одиночнорожденных, различий в когнитивном развитии обнаружено не было. Эти данные говорят о том, что некоторое снижение интеллекта у близнецов связано не столько с трудностями эмбрионального или родового периода, сколько с особенностями воспитания близнецов как пары. Во-первых, при рождении близнецов увеличивается нагрузка на семью, и у родителей остается гораздо меньше времени, которое они могут потратить на развивающие занятия с детьми. Во-вторых, растущие близнецы сами по себе являются социальной группой и испытывают меньше потребности в обмене информацией с внешним миром. Такая ситуация замкнутости близнецов друг на друга носит название "близнецовой ситуации" и часто приводит к отставанию близнецов в развитии. Так, у близнецов может развиваться особый, понятный только им автономный язык, развитие же полноценной речи, в результате, отстает. Это явление более характерно для МЗ близнецов, которые отличаются большей привязанностью друг к другу. Такое отставание в речевом развитии легко преодолевается, если детей на время разлучают (например, помещают в разные группы детского сада), и они бывают вынуждены общаться с другими детьми и взрослыми.
Таким образом, некоторые особенности развития близнецов приводят к тому, что близнецы как особая часть популяции несколько отличаются от остальной ее части. Эти отличия более отчетливо заметны в раннем, особенно младенческом, возрасте. Со временем некоторое отставание близнецов в физическом и умственном развитии компенсируется, и, по большей части, близнецы становятся сопоставимы с остальной популяцией.
Выводы
Существуют два типа близнецов - монозиготные и дизиготные (МЗ и ДЗ), или, что то же самое, - однояйцевые и двуяйцевые.
МЗ близнецы - это дети от многоплодной беременности, которые развиваются из одной оплодотворенной яйцеклетки (зиготы) в результате разделения одного зародыша на два самостоятельных организма на ранних стадиях эмбрионального развития. МЗ близнецы имеют идентичные генотипы (100% общих генов). МЗ близнецы всегда одного пола.
ДЗ близнецы - это дети от многоплодной беременности, которые развиваются из двух яйцеклеток, оплодотворенных двумя спермиями (из двух зигот). По своей генетической конституции ДЗ близнецы соответствуют обычным братьям и сестрам (сибсам), т.е. имеют в среднем 50% общих генов. ДЗ близнецы могут быть разного пола.
Частота рождения близнецов зависит от зиготности. В среднем ДЗ близнецы рождаются в два раза чаще, чем МЗ. Частота рождения ДЗ близнецов колеблется в зависимости от популяции.
Классический близнецовый метод предполагает сопоставление уровня сходства в парах МЗ и ДЗ близнецов и основан на следующих допущениях:
1) равенство сред для партнеров как в парах МЗ, так и в парах ДЗ близнецов;
2) отсутствие систематических различий между близнецами и неблизнецами;
3) отсутствие систематических различий между МЗ и ДЗ близнецами.
При чисто генетической детерминации индивидуальных различий (аддитивный эффект действия генов) величина корреляции между партнерами МЗ близнецовых пар должна приближаться к 1,0, а между близнецами ДЗ пар - к 0,5.
При средовой детерминации индивидуальных различий величина корреляций как МЗ, так и ДЗ близнецов должна приближаться к 1,0 (в силу общности среды).
Допущение о равенстве сред для близнецовых партнеров может нарушаться в силу следующих причин:
1) существуют специфические средовые условия, увеличивающие сходство МЗ близнецов;
2) существуют специфические средовые условия, уменьшающие сходство ДЗ близнецов;
3) существует ряд средовых условий, приводящих к неравноценности сред для партнеров как в парах МЗ, так и в прах ДЗ близнецов, что уменьшает их сходство.
Если изучаемая психологическая характеристика чувствительна к тем средовым условиям, равенство которых нарушается, показатель наследуемости, получаемый в близнецовом исследовании, может искажаться.
Помимо классического варианта, существует несколько разновидностей близнецового метода, в частности, метод разлученных близнецов, позволяющий наиболее четко разделить наследственные и средовые влияния.
в начало
Метод приемных детей
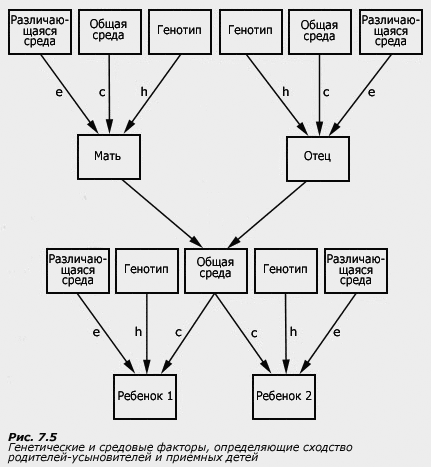
Исследование усыновленных детей является жесткой экспериментальной схемой, позволяющей четко развести влияние генетических и средовых факторов. Эта схема включает в себя, во-первых, сопоставление детей и их биологических родителей и, во-вторых, сопоставление детей и их родителей-усыновителей. Если дети были усыновлены в первые дни жизни и никогда не видели своих биологических родителей, то с ними они имеют только общие гены. С родителями-усыновителями, с которыми дети прожили всю жизнь, у детей нет никакого генетического сходства, но зато есть общие средовые условия:

В таблице отражен главный принцип работы метода приемных детей. Однако реальные коэффициенты корреляции, естественно, могут значительно отличаться от идеальных. Более высокая корреляция детей с биологическими родителями свидетельствует в пользу генотипических влияний, большая же корреляция с родителями-усыновителями - в пользу средовых. (Напомним, что эффекты внутриутробной среды у усыновленных детей определяются их биологическими матерями, поэтому, строго говоря, нельзя считать, что у таких детей нет элементов общей среды с их биологическими матерями. Если изучаемый признак чувствителен к влияниям внутриутробной среды, то в схемах исследования необходимо учитывать и материнский эффект.) Варианты этой схемы включают в себя, дополнительно к сопоставлению родителей и детей, еще и сравнение детей, не имеющих генетического сходства, но усыновленных в одну семью.
Поскольку отказ от своих детей и их последующее усыновление представляет собой явно не обыденную ситуацию, чистота метода приемных детей при исследовании психологических характеристик может подвергаться сомнению. Возможно ли набрать достаточно большую выборку детей, которые были усыновлены в первые дни жизни? Нет ли патологий у матерей, отказавшихся от своих детей (психических заболеваний, алкоголизма и т.д.), которые могут передаваться по наследству и через которые будут "преломляться" психологические особенности детей (что и приведет к сходству детей с их биологическими матерями)? Таких вопросов может быть много, и для того, чтобы не разбирать по одному разные сомнения по поводу метода приемных детей, рассмотрим, как было организовано одно из наиболее крупных исследований - Колорадское исследование приемных детей (DeFries J.C., Plomin R., Fulker D.W., 1994). Оно является хорошим доказательством того, что в исследовании приемных детей можно достаточно надежно развести генетические и средовые факторы, влияющие на изменчивость различных характеристик.
Прежде всего, выяснилось, что можно найти такую социальную группу, в которой психически нормальные и социально не деградировавшие женщины отказываются от своих детей. Ими оказались незамужние молодые женщины, в основном студентки, католического вероисповедания, которые по религиозным мотивам не могли избавиться от ребенка на первых стадиях беременности, но по разным причинам не могли или не хотели оставлять ребенка у себя (боялись осуждения родственников, не имели достаточно прочного материального положения, чтобы создать нормальные условия для ребенка, считали, что ребенку будет лучше в семье, где есть не только мать, но и отец и т.д.). Католическая община помогает таким женщинам найти семьи, которые хотят усыновить ребенка.
В таких случаях известен отец ребенка, и поэтому в исследовании могут быть получены и использованы данные не только матери, но и отца (социально-демографические параметры, показатели ряда психологических характеристик).
Для исследования были отобраны такие дети, которых усыновили в первый месяц жизни, при этом биологические матери проводили вместе с ребенком не более недели (в среднем 4 дня) и больше никогда с ним не встречались.
Социально-экономический статус биологических матерей, естественно, был ниже, чем статус семей-усыновителей (они были моложе, многие из них еще не получили профессию). Однако родители тех, кто отказался от детей - биологические бабушки и дедушки приемных детей - не отличались по социально-экономическому статусу от бабушек и дедушек в семьях-усыновителях. Биологические родители (и матери, и отцы) не отличались от родителей-усыновителей по своим интеллектуальным характеристикам, причем совпали не только средние показатели интеллекта, но и факторная структура измерявшихся переменных.
Корреляции между показателями интеллекта биологических родителей и родителей-усыновителей были близки к нулю. Незначимыми оказались и корреляции образования и профессионального статуса. Это говорит о том, что при усыновлении дети случайно распределялись по семьям: не было такого, чтобы ребенок более образованной биологической матери попадал в более образованную семью, а ребенок менее образованной матери - в менее образованную. Случайное распределение детей по семьям позволяет говорить о четком разделении в эксперименте генетических и средовых влияний.
Число приемных детей, которое было отобрано примерно за 10 лет, приближается к 300, т.е. выборка достаточно большая для того, чтобы выводы исследования могли быть надежными. Для повышения надежности данных в исследование включены также и контрольные семьи - обычные семьи, в которых сопоставляется сходство родителей и их собственных детей. Сравнение данных, полученных в приемных семьях, с данными, полученными в обычных семьях, позволяет проверить, не различаются ли стили родительско-детских отношений в двух типах семей.
В настоящее время дети, участвующие в Колорадском исследовании приемных детей, уже стали взрослыми, но исследование на этом этапе не оканчивается. Авторы предполагают проследить экспериментальную выборку до того времени, когда дети приблизятся к возрасту своих родителей, и их обследование можно будет провести теми же самыми психологическими методами.
Таким образом, схема исследования приемных детей, позволяющая надежно развести генетические и средовые факторы, влияющие на индивидуальные различия, может быть осуществлена в реальном эксперименте. Вместе с тем, многие исследователи считают, что метод приемных детей вряд ли будет играть существенную роль в будущих генетических работах, но связано это не с разрешающей способностью самого метода, а с традициями усыновления. Прежде всего, в ряде стран, и в частности в нашей, тайна усыновления охраняется законом: детям не принято сообщать о факте усыновления и, живя в приемной семье, они считают, что живут со своими биологическими родителями. В этих случаях проведение исследования приемных детей не представляется возможным. В тех странах, где факт усыновления не принято скрывать от детей, например, в США, где проведены наиболее многочисленные и чистые исследования приемных детей, постепенно меняется практика усыновления: все большее распространение получают такие варианты усыновления, при которых биологические родители ребенка, отдавая его в другую семью, не прекращают контакты с ним и принимают самое деятельное участие в его воспитании. Такая ситуация, несомненно, свидетельствует о либерализации взглядов в обществе (исчезает стигматизация матерей-одиночек, окружающие терпимее относятся к отдавшим своего ребенка в другую семью, если это не ущемляет интересов ребенка, и так далее). Однако такой вариант усыновления, если он станет общепринятым, сделает невозможным проведение генетических исследований, поскольку не позволит надежно развести генетические и средовые факторы, влияющие на изменчивость изучаемых характеристик.
Выводы
Метод приемных детей является жесткой экспериментальной схемой, позволяющей четко разводить влияния генетических и средовых факторов на изменчивость изучаемых психологических характеристик.
При использовании метода приемных детей производится сопоставление детей с их биологическими родителями и родителями-усыновителями.
Высокая корреляция между детьми и биологическими родителями свидетельствует о генотипических влияниях на изменчивость изучаемой характеристики. Высокая корреляция между детьми и родителями-усыновителями свидетельствует о средовых влияниях.
В ряде стран (в том числе и в России), в связи с существующими там традициями усыновления (тайна усыновления охраняется законом), применение метода приемных детей практически невозможно.
в начало
Семейные исследования
Анализ родословных
Метод анализа родословных первым начал применяться в генетике поведения. Им пользовался Ф. Гальтон для изучения наследственности таланта. Хотя этот метод как таковой в генетике поведения в настоящее время практически не применяется, что связано с его малой эффективностью, все же в генетике человека, и в особенности в медицинской генетике, метод изучения родословных является одним из основных. Поэтому коротко остановимся на описании этого метода.
Поскольку в генетике человека экспериментальные скрещивания невозможны, наследование того или иного признака изучают путем сбора данных в семьях. В семьях можно проследить те же закономерности менделевского расщепления и независимого распределения признаков, что и при экспериментальных скрещиваниях у растений и животных. В тех случаях, когда изучаются альтернативные (дискретные, качественные) признаки, анализ родословных помогает установить тип наследования (доминантный, рецессивный, сцепленный с полом). Через родословные можно получить сведения об аллелях и обнаружить сцепленные гены. В медицинской генетике метод родословных широко применяется при изучении наследования различных заболеваний или патологических отклонений. В современной психогенетике родословные необходимы для локализации генов на хромосомах при анализе сцепления, о котором пойдет речь в следующей теме.
Для большинства психологических признаков характерно полигенное наследование, поэтому метод родословных как таковой применять не имеет смысла. Однако применяемые здесь методы анализа внутрисемейного сходства (близнецы, приемные дети, родители, сибсы) можно считать расширенным вариантом метода родословных. Так, Ф. Гальтон в своем известном исследовании, описывая родословные знаменитых людей, рассматривал выдающиеся способности как качественный признак, вернее, как крайнюю степень выраженности количественного признака, при котором количество переходит в новое качество, которое можно назвать талантом.
Ф. Гальтон первым применил генеалогический метод для анализа наследования у человека, однако схемы родословных, составленные Ф. Гальтоном, внешне значительно отличаются от привычных для нас схем.
В современной генетике человека при составлении родословных пользуются системой специальных символов, которая была предложена в 1931 г.
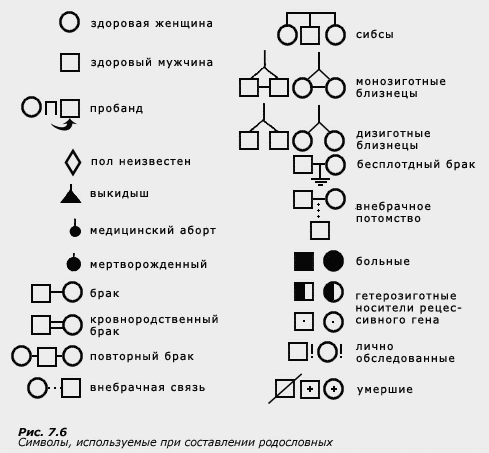
Носитель интересующего нас признака (например, пораженный болезнью или обладающий каким-либо талантом) называется пробандом. На схемах больные обозначаются зачерненными символами. Гетерозиготные носители рецессивного гена могут обозначаться символами, зачерненными наполовину. Поколения нумеруют сверху вниз римскими цифрами, а индивиды в пределах поколения нумеруются арабскими цифрами. Таким образом, каждый индивид в родословной имеет свой шифр (например, IV-7). Ф. Гальтон проанализировал множество родословных выдающихся людей и обнаружил, что частота родственных связей талантливых людей гораздо выше той, которую можно было бы ожидать при случайном распределении высокой одаренности. Вот как Ф. Гальтон описывает эту родословную. "Все семейство Бахов замечательно своим музыкальным талантом, который составлял принадлежность множества членов его и поддерживался в нем в восьми поколениях. Впервые он обнаружился в 1550 г., с особенной силой проявился в Иоганне Себастьяне (6-й по генеалогической таблице) и закончился Региной Сусанной, жившей еще в 1800 г. и находившейся в стесненных обстоятельствах. В этом семействе было более 20 выдающихся музыкантов. В сборнике биографий помещены жизнеописания по крайней мере 57 из его членов... По семейному обычаю, все члены этой семьи съезжались ежегодно, и в этих собраниях их единственным занятием была музыка. Около 1750 г. на таком собрании было не менее 120 Бахов".
Однако Ф. Гальтон вполне отдавал себе отчет в том, что не одна наследственность может быть причиной повышенной частоты таланта в некоторых семьях, особенно в семьях выдающихся государственных деятелей, ведь высокое положение отца в обществе открывает для его сына большие возможности, чем для сына обычного человека. Все же, на основании своих исследований Ф. Гальтон делает вывод о значительном влиянии наследственности на возникновение таланта и достижение известности.
После работ Ф. Гальтона неоднократно делались попытки подтвердить наследование гениальности. В качестве доказательств наследственной одаренности, помимо всем известного семейства Бахов, приводились примеры родословных различных знаменитых семейств. Представляет интерес, например, династия математиков Бернулли, которая в 6 поколениях дала 11 знаменитых ученых. 103 года представители этой семьи возглавляли кафедру математики в Базельском университете в Швейцарии. Но данные о семьях выдающихся людей - еще не доказательство наследственной передачи таланта, это лишь повод поставить об этом вопрос. Действительно, найдется немало примеров, когда выдающиеся музыканты или ученые были единственными в своем роду. Наблюдая семейную передачу какой-либо способности, нельзя с полной уверенностью утверждать, что она является наследственной, поскольку в семье помимо наследственной существует еще и культурная передача и корреляция между родителями и потомством может объясняться не только наследственностью, но и средой. И доказательство этому - большая встречаемость музыкальных семей в XVII-XVIII вв., чем в нынешнее время. Ведь в то время музыка была таким же ремеслом, как и кузнечное или портняжное, которое передавалось из поколения в поколения как средство обеспечения семьи.
Таким образом, высокая встречаемость выдающихся способностей у представителей одной семьи не является доказательством их наследуемости, поскольку генеалогический метод не позволяет разводить наследственные и средовые причины изменчивости количественных признаков. В настоящее время в генетике поведения генеалогический метод сам по себе не используется, его применяют в сочетании с другими (близнецовым, приемных детей, при анализе сцепления).
Исследования родственников в семьях
В семейных исследованиях рассматривается сходство членов одной семьи друг с другом. Сравниваемые родственники могут принадлежать к одному поколению. К их числу относятся братья и сестры (сибсы), родившиеся в одной семье и имеющие в среднем половину общих генов, а также родственники, имеющие меньшее генетическое сходство, - например, дети от разных браков - полусибсы (дети, имеющие одну и ту же мать, но разных отцов, или наоборот), двоюродные братья и сестры и т.д. Сравниваться могут и пары родственников, принадлежащие к разным поколениям: родители - с детьми, бабушки и дедушки - с внуками, тети и дяди - с племянниками.
Интерпретация результатов при таких сопоставлениях такая же, как и в близнецовом методе: о влиянии генотипа можно говорить в тех случаях, когда большей степени родства сопутствует и большее сходство (корреляция) по изучаемой психологической характеристике, например, если сибсы похожи больше, чем полусибсы; родители и дети - больше, чем бабушки и внуки, и т.д.
Семейное исследование, как и классический близнецовый метод, относится к "нежестким" экспериментальным схемам: как правило, родственники, имеющие больше общих генов, имеют и более похожие условия жизни и больше возможностей влиять друг на друга, т.е. генетические и средовые условия, влияющие на сходство родственников, оказываются не независимыми друг от друга.
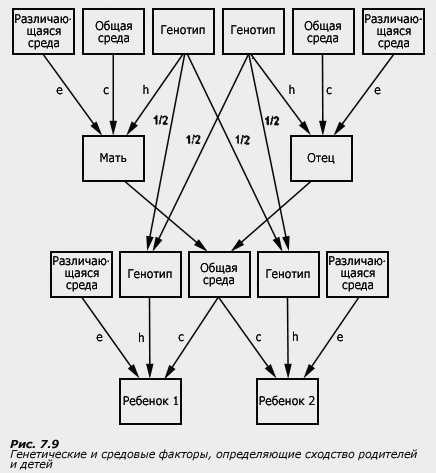
В связи с этим, в рамках экспериментальной схемы семейного исследования более надежным для определения роли генотипа и среды является сопоставление двоюродных братьев и сестер с троюродными (поскольку и те и другие, по крайней мере, не живут обычно под одной крышей), чем родных братьев и сестер с двоюродными.
Еще одна проблема возникает при сравнении родственников, принадлежащих к разным поколениям, например, родителей и детей. Во-первых, влияние генотипа на некоторые психологические особенности может изменяться с возрастом. Во-вторых, методы диагностики психологических особенностей, рассчитанные на людей разного возраста, могут быть не вполне сопоставимы. В связи с этим роль генотипических факторов в исследованиях родителей и детей может занижаться: дети могут оказаться непохожими на родителей по какой-либо характеристике, вариативность которой в значительной степени связана с генотипом. (Напомним, что в семейных исследованиях мерой сходства является величина корреляции и речь о сходстве в абсолютных величинах не идет).
Выходом из этой ситуации может быть только проведение долговременных исследований: обследовать родителей по какой-либо характеристике, подождать, пока их дети вырастут до взрослого возраста (того, в котором обследовались их родители), и повторить исследование на детях.
Выводы
Для генетического анализа дискретных признаков может применяться метод изучения родословных. В случае менделирующих признаков анализ родословных позволяет определить тип наследования (аутосомный, сцепленный с полом, доминантный, рецессивный и т.п.).
При изучении количественных признаков можно проводить семейные исследования - рассматривается сходство членов одной семьи друг с другом.
О влиянии генотипа говорят в тех случаях, когда большей степени родства соответствует большее сходство (корреляция) по изучаемой характеристике.
Семейное исследование в генетике поведения не позволяет четко развести средовые и генетические влияния и относится к "нежестким" экспериментальным схемам, поскольку родственники, имеющие больше общих генов, имеют и более похожие условия среды.
К сложностям семейных исследований можно отнести различия в возрасте между родственниками, принадлежащими к разным поколениям.
в начало
Сопоставление результатов, полученных разными методами
Одним из способов проверки надежности результатов, получаемых при исследовании роли генотипа и среды в разных психологических характеристиках, является сопоставление результатов, полученных разными методами.
Допустим, при исследовании одной и той же психологической характеристики близнецовым и семейным методами, а также методом приемных детей были получены такие результаты, какие приведены в примере 1:
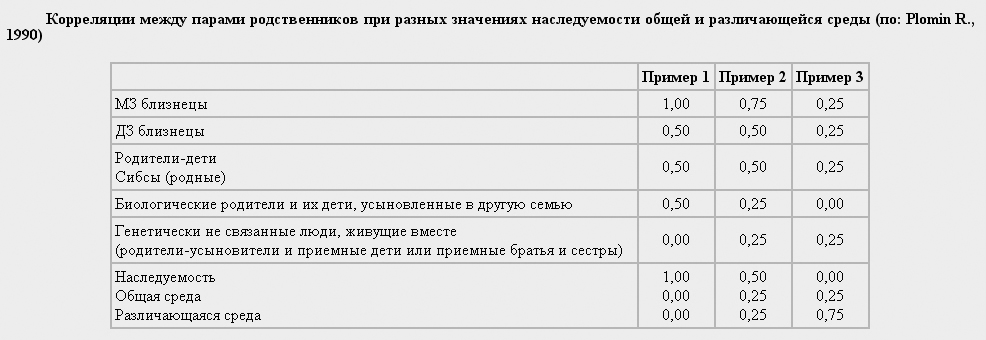
Эти результаты свидетельствуют о том, что все индивидуальные различия по изучаемой характеристике определяются генотипом: МЗ близнецы оказываются полностью идентичны по этой характеристике, остальные родственники похожи в той степени, которая соответствует их генетическому сходству (корреляции родителей и детей равны 0,5; корреляции сибсов равны 0,5; корреляции родителей и приемных детей равны 0,0).
Во втором примере корреляции между родственниками определяются на 50% наследуемостью и на 25% - общей средой, поэтому сходство между МЗ близнецами будет составлять 50% от их генетического сходства (напомним, что у МЗ близнецов генетическое сходство составляет 100%) плюс сходство за счет общей среды (25%), т.е. в результате корреляция между МЗ близнецами составит 0,75. Сходство между ДЗ близнецами будет складываться таким же образом: поскольку генетическое сходство ДЗ близнецов составляет 50%, при 50-процентной наследуемости мы будем иметь 25% сходства за счет общих генов и 25% - за счет общей среды, соответственно корреляция для ДЗ близнецов составит 0,5. Сходство приемных детей и усыновивших их родителей будет определяться только общей средой (25%), поэтому их корреляция составит 0,25.
В третьем примере сходство между родственниками определяется только средовыми параметрами (25% за счет общей среды), поэтому для тех родственников, которые имеют общую среду, корреляция будет равна 0,25.
Все три приведенных примера демонстрируют результаты, которые получаются в тех случаях, когда данные, полученные разными методами, не вступают в противоречие. Если же результаты исследования одной и той же психологической характеристики, полученной разными методами, не совпадут, то можно выяснить причины, изменяющие внутрипарное сходство тех или иных родственников. Например, если сходство ДЗ близнецов превышает сходство сибсов, можно предположить влияние специфических близнецовых эффектов, имеющих средовое происхождение и увеличивающих сходство ДЗ близнецов.
Выводы
Близнецы и близнецовый метод
Существуют два типа близнецов - монозиготные и дизиготные (МЗ и ДЗ), или, что то же самое, - однояйцевые и двуяйцевые.
МЗ близнецы - это дети от многоплодной беременности, которые развиваются из одной оплодотворенной яйцеклетки (зиготы) в результате разделения одного зародыша на два самостоятельных организма на ранних стадиях эмбрионального развития. МЗ близнецы имеют идентичные генотипы (100% общих генов). МЗ близнецы всегда одного пола.
ДЗ близнецы - это дети от многоплодной беременности, которые развиваются из двух яйцеклеток, оплодотворенных двумя спермиями (из двух зигот). По своей генетической конституции ДЗ близнецы соответствуют обычным братьям и сестрам (сибсам), т.е. имеют в среднем 50% общих генов. ДЗ близнецы могут быть разного пола.
Частота рождения близнецов зависит от зиготности. В среднем ДЗ близнецы рождаются в два раза чаще, чем МЗ. Частота рождения ДЗ близнецов колеблется в зависимости от популяции.
Классический близнецовый метод предполагает сопоставление уровня сходства в парах МЗ и ДЗ близнецов и основан на следующих допущениях:
1) равенство сред для партнеров как в парах МЗ, так и в парах ДЗ близнецов;
2) отсутствие систематических различий между близнецами и неблизнецами;
3) отсутствие систематических различий между МЗ и ДЗ близнецами.
При чисто генетической детерминации индивидуальных различий (аддитивный эффект действия генов) величина корреляции между партнерами МЗ близнецовых пар должна приближаться к 1,0, а между близнецами ДЗ пар - к 0,5.
При средовой детерминации индивидуальных различий величина корреляций как МЗ, так и ДЗ близнецов должна приближаться к 1,0 (в силу общности среды).
Допущение о равенстве сред для близнецовых партнеров может нарушаться в силу следующих причин:
1) существуют специфические средовые условия, увеличивающие сходство МЗ близнецов;
2) существуют специфические средовые условия, уменьшающие сходство ДЗ близнецов;
3) существует ряд средовых условий, приводящих к неравноценности сред для партнеров как в парах МЗ, так и в прах ДЗ близнецов, что уменьшает их сходство.
Если изучаемая психологическая характеристика чувствительна к тем средовым условиям, равенство которых нарушается, показатель наследуемости, получаемый в близнецовом исследовании, может искажаться.
Помимо классического варианта, существует несколько разновидностей близнецового метода, в частности, метод разлученных близнецов, позволяющий наиболее четко разделить наследственные и средовые влияния.
Метод приемных детей
Метод приемных детей является жесткой экспериментальной схемой, позволяющей четко разводить влияния генетических и средовых факторов на изменчивость изучаемых психологических характеристик.
При использовании метода приемных детей производится сопоставление детей с их биологическими родителями и родителями-усыновителями.
Высокая корреляция между детьми и биологическими родителями свидетельствует о генотипических влияниях на изменчивость изучаемой характеристики. Высокая корреляция между детьми и родителями-усыновителями свидетельствует о средовых влияниях.
В ряде стран (в том числе и в России), в связи с существующими там традициями усыновления (тайна усыновления охраняется законом), применение метода приемных детей практически невозможно.
Семейные исследования
Для генетического анализа дискретных признаков может применяться метод изучения родословных. В случае менделирующих признаков анализ родословных позволяет определить тип наследования (аутосомный, сцепленный с полом, доминантный, рецессивный и т.п.).
При изучении количественных признаков можно проводить семейные исследования - сопоставлять сходство друг с другом членов одной семьи.
О влиянии генотипа говорят в тех случаях, когда большей степени родства соответствует большее сходство (корреляция) по изучаемой характеристике.
Семейное исследование в генетике поведения не позволяет четко развести средовые и генетические влияния и относится к "нежестким" экспериментальным схемам, поскольку родственники, имеющие больше общих генов, имеют и более похожие условия среды.
К сложностям семейных исследований можно отнести различия в возрасте между родственниками, принадлежащими к разным поколениям.
Сопоставление результатов, полученных разными методами
Сопоставление результатов исследования одной и той же психологической характеристики с применением разных методов - близнецового, семейного и метода приемных детей - позволяет оценивать их надежность.
Если результаты исследования одной и той же психологической характеристики, полученные разными методами, вступают в противоречие, необходим дополнительный анализ причин, влияющих на внутрипарное сходство тех или иных родственников.
в начало
Методы, использующие молекулярно-генетические технологии и моделирование на животных
Методы психогенетики, описанные в предыдущей теме 7, не могут быть использованы для поиска генов, имеющих отношение к детерминации поведения. Они лишь позволяют статистически оценить наследственные и средовые эффекты, определяющие изменчивость психологических характеристик в конкретных популяциях людей. Получаемые при этом оценки наследуемости и средовых эффектов общей и индивидуальной среды оказываются малоинформативными в плане познания механизмов действия наследственности и среды на формирование конкретных психологических особенностей людей.
Новая эра в психогенетике началась с момента основания в конце 80-х гг. долгосрочной программы "Геном человека", о которой мы уже упоминали. Широкомасштабные исследования, проводимые в последнее время в рамках проекта "Геном человека", открыли новые возможности для исследований в генетике поведения. Можно с уверенностью сказать, что за последние несколько лет кардинально изменились акценты в генетике поведения. Приход молекулярно генетических технологий в генетику поведения человека можно считать революционным этапом, в корне преобразовавшим эту область.
Прежде чем начать описание конкретных методических приемов, используемых для поиска генов и изучения механизмов их действия, необходимо коротко остановиться на основных достижениях молекулярной генетики последних лет, чтобы понять, на чем основываются новые технологии.
в начало
Геномика и психогенетика
Геномика, пожалуй, самая молодая область генетики. Длительное время понятие "геном" определяли как совокупность генов в гаплоидном наборе хромосом определенного биологического вида. Когда говорили о геноме человека, то имелся в виду гаплоидный набор хромосом вида "Homo sapiens" с содержащимися в нем генами. Сейчас положение кардинально изменилось.
Во-первых, были накоплены сведения о внехромосомной ДНК, находящейся в цитоплазме и ядрах клеток вне хромосом.
Во-вторых, оказалось, что ДНК - это не просто цепочка генов, следующих один за другим: структура и функции ее намного сложнее. Структурные гены, т.е. те, которые кодируют полипептидную цепь или молекулу РНК, занимают лишь небольшую часть ДНК (примерно 3-5%). Существует огромное количество межгенных участков с пока еще неясными функциями, образующих вместе с генами всю совокупность нуклеотидных последовательностей ДНК. В настоящее время геномом называется полный состав ДНК клетки.
Геномика изучает принципы построения геномов и их структурно-функциональную организацию. Геномика еще очень молода, но столь стремительно развивается, что в ее рамках уже появились самостоятельные направления: структурная геномика, функциональная геномика, медицинская геномика, компьютерная геномика, сравнительная геномика, эволюционная геномика, этническая геномика. Структурная геномика изучает нуклеотидные последовательности ДНК, в том числе строение и локализацию генов, определяет их границы, изучает строение межгенных участков, обеспечивающих регуляторные функции ДНК. В задачу структурной геномики входит построение генетических карт организма. Функциональная геномика решает вопросы, связанные с идентификацией функций отдельных участков генома, в том числе и структурных генов, а также механизмы их взаимодействия в клеточном ансамбле. С функциональной геномикой тесно связана еще одна молодая область молекулярной биологии - протеомика, изучающая функционирование белков в клетках. Геномика не может развиваться без современных компьютерных технологий, поскольку оперирует огромными базами данных (компьютерная геномика). Достижения структурной и функциональной геномики применяются в прикладных целях (медицинская геномика, сельскохозяйственная геномика). Сравнительная и эволюционная геномика изучает сходства и различия геномов разных организмов и происхождение генетического полиморфизма, свойственного всем живым организмам на Земле. Этническая геномика изучает особенности геномного полиморфизма отдельных популяций, рас, народностей.
Современные представления о геноме человека сложились благодаря нескольким открытиям молекулярной биологии (в 70-х гг.х ХХ столетия), позволившим получать неограниченные количества ДНК (открытие полимеразной цепной реакции – ПЦР) и расшифровывать последовательности нуклеотидов в ее цепях (секвенирование). Немаловажным фактором в стремительном прогрессе геномики была автоматизация процессов секвенирования и развитие информационных технологий. В США и Великобритании разработаны роботы-секвенаторы, которые были названы геномотронами. Эти приборы осуществляют до 100 000 полимеразных реакций в час. В течение недели секвенаторы могут осуществить расшифровку нуклеотидных последовательностей длиной в несколько миллионов пар нуклеотидов. В результате создания высоких биотехнологий стало возможным в обозримые сроки просеквенировать весь геном человека, что и было выполнено примерно за 10 лет в результате действия международной и национальных программ проекта "Геном человека" (1990-2000).
Результатом этих работ стало получение информации не только о последовательности нуклеотидов всей ДНК человека, но и открытие громадного количества новых полиморфизмов - генетических вариантов последовательностей нуклеотидов одного и того же участка ДНК у разных людей.
Когда мы в теме 3 на примере групп крови системы АВ0 вводили понятие множественного аллелизма, мы фактически разбирали случай генетического полиморфизма у человека по определенным белкам крови. Аллели - это различные формы гена (отсюда - полиморфизм). Геномный полиморфизм представляет собой участки ДНК (функции которых могут быть в настоящее время неизвестны), варьирующие у отдельных индивидов. Любые изменения в ДНК ведут к возникновению генетического полиморфизма. Полиморфизм может возникать в результате замены или потери нуклеотидов. Это однонуклеотидный полиморфизм.
Полиморфизм может быть также результатом количественных изменений генома. В этом случае варьирует число нуклеотидных повторов. Полиморфизмы наблюдаются как в смысловых (экзоны), так и в несмысловых (интроны) участках генома. Однонуклеотидные полиморфизмы составляют основу генетической вариативности. Они встречаются по всему геному и отражают прошлые мутации. Вся длина генома составляет 3,2 млрд пар нуклеотидов. К 2001 г. идентифицировано и картировано 1,42 млн однонуклеотидных полиморфизмов. По расчетам генетиков, люди оказываются идентичными по нуклеотидным последовательностям на 99,9%. Таким образом, столь заметная вариативность между людьми возникает за счет различий лишь в 1% генома (Бочков Н.П., 2002). Данные секвенирования показали, что в геноме человека чуть более 30 000 генов, тогда как раньше считалось, что их, по крайней мере, в два-три раза больше. Большинство генов имеют размеры до 50 000 пар нуклеотидов. Число синтезируемых белковых продуктов превышает число генов в 1,5-2 раза. Это является результатом альтернативного сплайсинга при вырезании интронов (рис.8.2. a, b, c). Как размеры генов, так и число экзонов и интронов в них варьирует в довольно широких пределах:
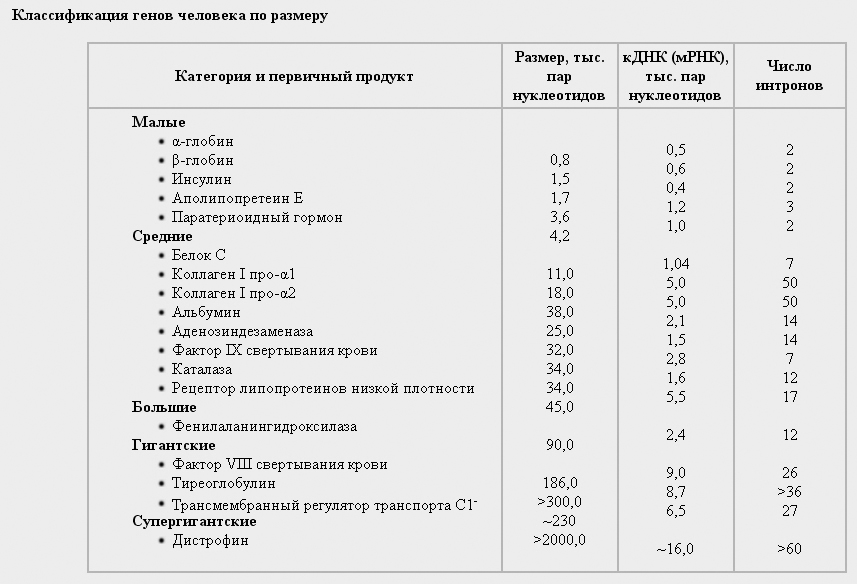

Благодаря открытию геномного полиморфизма генетики получили широкие возможности для картирования генов, т.е. локализации генов на хромосомах. Распределенные по всему геному, полиморфизмы используются как генетические маркеры. Генетическими маркерами называются участки ДНК с известной локализацией и множественными аллельными формами. Они служат как бы опорными точками для картирования генов (подробнее об этом см. в следующем параграфе). Поиск генетических маркеров был одной из самостоятельных задач проекта "Геном человека". Концептуальная модель использования маркеров для локализации генов чрезвычайно проста. Если два индивида, связанные кровным родством, имеют идентичные маркеры, высока вероятность того, что рядом расположенные участки ДНК, в силу тесного сцепления, будут у них также идентичны. Если у этих родственников наблюдается к тому же одно и то же заболевание, т.е. они являются по нему конкордантными, можно предполагать, что ген болезни расположен рядом с маркером. Похожий принцип лежит и в основе картирования локусов количественных признаков (Quantitative Trait Loci - QTL), которые до недавнего времени, в отличие от признаков качественных, практически не могли быть картированы.
Основные молекулярные методы, разработанные на основе исследований генома человека и животных, были ассимилированы генетикой поведения и в настоящее время широко используются для локализации генов, связанных с различными психическими заболеваниями и отклонениями в развитии психики. Наиболее успешно решается проблема локализации генов моногенных заболеваний (фенилкетонурия, хорея Гентингтона), наследуемых по Менделю. Гораздо труднее идентифицировать гены, имеющие отношение к мультифакториальным заболеваниям (шизофрения, депрессия, алкоголизм и т.п.) или нормально варьирующим признакам (интеллект, темперамент, особенности личности).
Можно выделить три основных экспериментальных подхода, использующих молекулярные технологии при работе с людьми. Это различные варианты метода анализа сцепления, анализ ассоциаций и непосредственный анализ ДНК, т.е. определение последовательности нуклеотидов (секвенирование) и идентификация мутаций. Кроме исследований, ведущихся на человеке, современная психогенетика привлекает также методы моделирования на животных. Остановимся на каждом из этих подходов более подробно.
в начало
Анализ сцепления
Классический вариант анализа сцепления
Гены в хромосомах располагаются линейно, один за другим. Этим обстоятельством определяются две основные особенности наследования генов, относящихся к одной хромосоме. Во-первых, такие гены имеют тенденцию передаваться совместно, поэтому их называют сцепленными. Во-вторых, хотя эти гены в процессе кроссинговера и могут рекомбинировать, их рекомбинация не является полностью случайной: вероятность совместной передачи повышается с уменьшением расстояния между генами.
Известный американский генетик Томас Морган еще в начале ХХ века предложил использовать явление сцепления для локализации генов, расположенных на одной хромосоме. Этот метод генетического анализа получил название анализ сцепления, а результатом его применения стало создание генетических карт, т.е. схем расположения конкретных генов на конкретных хромосомах. Генетические карты животных и растений, для которых возможны экспериментальные скрещивания, создаются постепенно в процессе наблюдений за частотой рекомбинации (чем чаще гены передаются совместно, тем ближе они расположены на хромосоме). Используя рекомбинантный анализ, Т. Морган и его коллеги успешно локализовали многие гены плодовой мушки дрозофилы. В настоящее время созданы подробные генетические карты для многих видов экспериментальных организмов и ряда сельскохозяйственных культур.
У человека экспериментальные скрещивания невозможны, поэтому классический способ рекомбинантного анализа может применяться весьма ограниченно, лишь на основе информации, которую дает анализ родословных. Чтобы выявить кроссинговер у человека, нужно располагать либо большой родословной, либо иметь несколько небольших родословных. Проще всего по родословной обнаруживается Х-сцепленное наследование, т.е. расположение генов в Х-хромосоме, поскольку характерный признак или заболевание наблюдается у лиц одного пола. Однако чтобы установить расположение гена в конкретном сегменте Х-хромосомы, нужны дополнительные схемы анализа, требующие применения новейших методик. Почти все Х-сцепленные гены (а их более 100) были картированы с помощью анализа родословных.
Для того чтобы картировать гены, принадлежащие аутосомам, необходимо сначала выявить группы генов, относящихся к определенной хромосоме. Простой анализ родословных не позволяет это сделать. Впервые локализация гена в аутосоме оказалась возможной благодаря наличию морфологического маркера хромосомы. Такой маркер в виде вторичной перетяжки был обнаружен на длинном плече первой хромосомы человека. Оказалось, что примерно в 0,5% случаев в популяции эта морфологическая особенность первой хромосомы отличается от нормы: перетяжка оказывается тоньше и длиннее. Было обнаружено, что такой вариант перетяжки наследуется доминантно. С этим морфологическим маркером оказался тесно сцепленным локус группы крови Даффи, а еще ранее было показано, что ген Даффи сцеплен с врожденной очаговой катарактой. Помимо визуально наблюдаемых морфологических особенностей хромосом, локализации генов на конкретных хромосомах могут способствовать анализ делеций (выпадений участков хромосомы) и некоторых других хромосомных аномалий, которые влекут за собой изменение ферментативной активности.
Таким образом, чтобы локализовать ген на определенной хромосоме, необходим маркер, по отношению к которому можно оценивать тесноту сцепления. Такими маркерами могут быть гены, локализация которых в данной хромосоме уже известна (например, локализация генов цветовой слепоты в Х-хромосоме), а также различные морфологические маркеры. В последнее время, благодаря успехам генной инженерии, стало возможным выявление молекулярных маркеров в ДНК человека. Огромные возможности для локализации генов появились с разработкой новых методов гибридизации клеток (например, клеток человека и мыши). В результате число уже локализованных генов человека растет с поразительной быстротой. Как показывают расчеты, весь геном человека может быть картирован, если исследователи будут располагать всего несколькими сотнями маркеров, которые случайно распределены по хромосомам (Фогель Ф., Мотульски А., 1989). Открытие множества геномных полиморфизмов человека обогатило генетику человека сотнями новых маркеров, благодаря которым проблема локализации генов и картирования хромосом успешно решается. Секвенирование генома позволяет составить полные генетические карты для всех хромосом. Карты хромосом постоянно пополняются новыми мутациями. В настоящее время наиболее полная генетическая карта составлена для генома митохондрий.
Мерой "генетического расстояния" служит частота рекомбинации между маркером и исследуемым геном. Это расстояние измеряется в морганидах или сантиморганидах (сМ), единицах, названных так в честь Томаса Моргана. Одна сантиморганида соответствует расстоянию между генами, рекомбинация между которыми происходит с частотой 1%. Карты генома могут составляться в двух вариантах. Одни из них - карты генетического сцепления - строятся на основе анализа данных по наследованию гена или маркера в ряду поколений, другие - физические карты - на прямом исследовании носителей генетической информации (Пузырев В.П., Степанов В.А., 1997). Генетическое расстояние в 1 сМ примерно равно физическому расстоянию в 1 миллион пар оснований. Длина генетической карты генома человека составляет около 3000-3500 сантиморганид (Бочков Н.П., 2002).
Если частота рекомбинации соответствует 50%, это означает, что два локуса находятся на таком расстоянии, что наследуются независимо. Если два гена передаются совместно в 10 поколениях, вероятность того, что это происходит случайно, то есть гены не являются сцепленными, ничтожно мала (1/1000 или 1/103). В научной практике эта вероятность выражается в логарифмической шкале (LOD score) и составляет в данном случае 3 lod. При работе с родословными пользуются различными таблицами лод-баллов и специально разработанными алгоритмами. Благоприятные возможности для определения сцепления по родословной существуют, если один из родителей является гетерозиготным по двум различным генам, а второй - гомозиготным. Чаще всего анализ сцепления проводится для часто встречающихся в популяции маркеров или в тех случаях, когда часто встречающийся маркер сопутствует редкому наследственному заболеванию. Как доказательство сцепления обычно рассматривается лод-балл не менее 3 (1000:1 в пользу сцепления). В настоящее время разработаны специальные компьютерные программы для анализа сцепления по родословным. Широко используется программа LIPED, которая позволяет получать оценки максимального правдоподобия параметров сцепления по данным родословных (Ott, 1991), а также программа LINCAGE (Lathrop et al., 1984).
Анализ сцепления чаще всего используется при изучении альтернативных (качественных) признаков. Делаются попытки применять этот метод и для анализа количественных признаков в расчете на то, что удастся обнаружить главные гены, влияющие на изучаемые характеристики. Теоретически такой подход вполне оправдан, однако, как отмечают Ф. Фогель и А. Мотульски (1989), следует соблюдать большую осторожность в интерпретации данных. Во-первых, при анализе большого числа количественных признаков существует риск обнаружить сцепление за счет чисто случайных флуктуаций критериев значимости. Во-вторых, корреляция, наблюдаемая в семьях, может возникать не только за счет сцепления, но и за счет наличия ассортативности по исследуемому признаку. Следует отметить, что пока анализ сцепления для количественных признаков испытывает значительные трудности и часто не приносит ожидаемых результатов, однако с появлением молекулярных маркеров ДНК возможности анализа и интерпретации данных постоянно возрастают.
Картирование локусов количественных признаков (ЛКП)
Описанный выше метод анализа сцепления в классическом варианте, как правило, используется для альтернативных признаков, наследуемых по Менделю. Для количественных признаков разработана несколько отличающаяся модель картирования - это так называемый метод QTL (Quantitative Trait Loci), или картирование ЛКП (локусов количественных признаков). ЛКП представляют собой полигенные системы, обеспечивающие непрерывную вариативность признака в популяции.
Метод картирования ЛКП основывается на исследовании ДНК у пар близких родственников, чаще всего сибсов. Идея его очень проста. Мы уже знаем, что сибсы имеют в среднем примерно 50% общих аллелей. В каждой конкретной паре сибсов совпадающие аллели будут отличаться. Таким образом, у детей одних и тех же родителей в данном локусе аллели матери и отца могут представлять различные сочетания:
Рассмотрим ген, который имеет четыре аллеля: А, В, С, D. Допустим, мать обладает аллелями А и В, а отец - аллелями С и D. Каждый ребенок может получить от матери один из ее аллелей (А или В) и от отца - один из его аллелей (С или D), при этом ребенок наследует свой генотип независимо от других детей в этой семье. Соответственно возможны четыре различных сочетания. Сравнение генотипов, представленных на рисунке, показывает, что любой ребенок в этой семье, унаследовавший, к примеру, генотип АС, может иметь от 0 до 2 общих аллелей с любым из своих братьев и сестер. Предположим, мы выявили пару сибсов, которые имеют два общих аллеля. Эти сибсы оказались более похожими по какому-либо количественному признаку, чем сибсы, имеющие один общий аллель. Последние, в свою очередь, обнаружили большее сходство, чем те сибсы, у которых общих аллелей в этом локусе не было совсем. На основании этих наблюдений, мы можем предполагать, что этот ген влияет на интересующий нас количественный признак. Если разница в количестве совпадающих аллелей никак не отражается на количественных соотношениях признака у сибсов, значит тестированный ген не имеет отношения к изучаемому признаку. Обычно при таком подходе вместо реальных генов используются маркеры и вычисляются корреляции между наличием маркера и степенью выраженности количественного признака. При анализе группы "количественных локусов" корреляции вычисляются для каждого из нескольких маркеров, расположенных на определенном участке хромосомы. Величина коэффициентов меняется от локуса к локусу. На основании дальнейшего анализа определяют участок предполагаемой локализации гена, вносящего свой вклад в количественный признак.
В некотором смысле метод картирования ЛКП с использованием пар сибсов напоминает близнецовый метод: сибсы, у которых совпадают оба аллеля аналогичны МЗ близнецам (100% общих генов). Сибсы, у которых совпадает 1 аллель аналогичны ДЗ близнецам (50% общих генов). Сибсы, не имеющие совпадений, аналогичны неродственникам (0% общих генов).
Целью картирования ЛКП является поиск не единичного гена, определяющего сложный признак, а скорее множественных генов, принимающих участие в формировании популяционной вариативности по изучаемой характеристике. Метод картирования ЛКП достаточно эффективен для выявления действия так называемых главных генов, т.е. генов, вносящих основной вклад в формирование количественного признака.
Предположим, имеется ген, участие которого определяет 10% вариативности признака, кроме того, существует еще 5 генов, каждый из которых дает вклад в 2%, и еще 10 других генов с вкладом в изменчивость в 1%. Если эффекты этих ЛКП (или QTL) независимы, то все они вместе будут определять 30% вариативности (10 + 2•5 + 1•10 = 30). Для генов, наследуемых по Менделю и имеющих вклад, оцениваемый в 100%, эффект порядка 10% может показаться весьма малым. Однако для различных психических расстройств такой вклад считается достаточно большим. На сегодняшний день метод картирования ЛКП используется для изучения количественных признаков или заболеваний с наследственной предрасположенностью и обладает разрешающей способностью, позволяющей выявлять гены с эффектом порядка 10% (Plomin R., McGuffin P., 2003). Малые эффекты (порядка 1%) пока недоступны для обнаружения. Несомненно, что поиск генов комплексных заболеваний и других мультифакториальных признаков представляет собой весьма сложную задачу.
Среди различных подходов, используемых в методе картирования ЛКП, наиболее популярным является анализ сибсовых пар (sibpair QTL linkage analysis), принцип которого разобран выше. Наиболее часто используемый вариант этого метода - модель пораженной пары (affected sibpair design). При такой схеме исследования оба сибсовых партнера имеют одинаковый диагноз или крайнее выражение какого-либо количественного признака. В среднем у сибсов должно наблюдаться совпадение в одном из двух аллелей (50%). Связь тестируемого участка хромосомы с болезнью можно предполагать, если у пораженных болезнью (т.е. конкордантных) сибсов совпадение аллелей будет наблюдаться в гораздо большем проценте случаев, чем ожидаемые 50%.
Анализ ассоциаций. Метод гена-кандидата
Еще один подход к картированию ЛКП использует принцип неравновесного сцепления (linkage disequilibrium) (или ассоциации). Под неравновесным сцеплением понимаются конкретные комбинации аллелей, по которым наблюдается неслучайное распределение в исследуемой группе лиц по сравнению с общей популяцией. В связи с этим частоты этих аллелей в популяции не соответствуют ожидаемым по случайному закону. Метод анализа ассоциаций может обнаруживать ЛКП, которые отвечают за гораздо меньший процент вариативности, чем позволяет анализ обычного сцепления. Ассоциация есть корреляция между отдельными аллелями и признаками в популяции. Например, известен ген, который имеет название аполипопротеин Е (АРОЕ). Этот ген имеет аллель АРОЕ-4, частота которого у людей, пораженных болезнью Альцгеймера (с поздним проявлением) составляет 40%, в то время как в контрольной группе - всего 15%.
Болезнь Альцгеймера - наследственное заболевание человека, которое характеризуется прогрессирующим нарушением памяти, утратой навыков, психотическими расстройствами, гиперкинезами и т.п. Связано с диффузной атрофией мозга. Проявляется в пожилом возрасте.
АРОЕ-4 обладает значительным эффектом, но наличие этого аллеля не является решающим фактором в развитии заболевания. Оно лишь повышает его риск. При этом около трети пациентов с болезнью Альцгеймера не имеют в своем генотипе АРОЕ-4, а примерно половина тех людей, что им обладают, доживают до 80 лет, не проявляя признаков болезни (Williams, 2003; цит. по: Plomin & McGuffin, 2003).
В большинстве исследований, использующих метод ассоциаций, сначала формулируется соответствующая гипотеза о влиянии какого-либо известного гена на поведенческий признак или заболевание. Этот ген называется геном-кандидатом (candidate gene). Так, в терапии депрессии и некоторых других психических заболеваний, связанных с повышенным беспокойством и тревогой, широко используются препараты, действующие на белок-переносчик серотонина в нервной системе. Этот белок является важным элементом связи между нейронами, продуцирующими серотонин. Другие исследования подтверждают, что обмен серотонина в нервной системе имеет отношение к развитию состояний тревоги и депрессии. Соответственно, ген, кодирующий белок-переносчик серотонина, рассматривается как ген-кандидат. Далее гипотеза проверяется с использованием анализа сцепления, метода ассоциаций и мутационного анализа.
Вообще гены, кодирующие белки, участвующие в процессах передачи информации в нервной системе, вызывают повышенный интерес исследователей как возможные гены-кандидаты для поведенческих признаков. Например, геном-кандидатом для поведения, связанного с рисками (поиск новизны - novelty seeking), является ген, кодирующий рецептор дофамина.
В ассоциативных исследованиях сопоставляется встречаемость определенного аллеля гена-кандидата в популяции в целом и у лиц, обладающих/не обладающих изучаемым типом поведения или пораженных болезнью. Если данный аллель чаще встречается у людей с выраженным поведенческим признаком (или заболеванием), чем у тех, кому оно совершенно не свойственно (или здоровых), то можно предполагать, что аллель играет определенную роль в наследственной детерминации признака. Однако полностью полагаться на результаты не следует, поскольку может случиться так, что ген-кандидат не сам регулирует поведение, а просто тесно сцеплен с тем геном, который на самом деле имеет отношение к признаку. В этом смысле на исследователях лежит определенная ответственность, поскольку они должны учесть все альтернативные возможности интерпретации получаемых результатов и только после этого делать заключение об ассоциации гена и признака.
в начало
Прямой анализ ДНК и выявление мутаций
Когда в результате анализа сцепления предполагаемое место гена на хромосоме определено, это означает всего лишь, что мы знаем, где находится данный ген, но этого недостаточно для того, чтобы понять механизмы изучаемого заболевания или типа поведения. Главная задача исследователей - не установить, где расположен ген, а понять, как он работает, и как факторы среды могут влиять на формирование изучаемой характеристики. До недавнего времени такая задача вообще не имела решения в рамках тех генетико-эпидемиологических подходов, которыми пользовалась психогенетика (см. предыдущую тему). Сейчас положение в корне изменилось: современные молекулярно-генетические методы позволяют непосредственно изучать тот участок молекулы ДНК, в котором расположен ген, связанный с контролем поведенческого признака. Гораздо меньшие возможности существуют пока для того, чтобы изучать, как среда влияет на работу гена.
Сейчас коротко опишем некоторые методы, которые применяются при непосредственном анализе ДНК. Вначале рассмотрим некоторые основные достижения генетики последних лет, которые создали предпосылки для массового использования молекулярно-генетических методов.
Осуществление проекта "Геном человека" было бы невозможно без применения методов генетической инженерии. Это направление в генетике возникло в начале 70-х гг. В его основе лежит методология конструирования и получения рекомбинантных ДНК. Рекомбинантными называются молекулы ДНК, которые получаются путем объединения in vitro (в пробирке) чужеродных фрагментов ДНК, которые в природе не встречаются. При этом применяются методы обратной транскрипции, позволяющие с помощью особого фермента (обратной транскриптазы) на основе фрагментов молекулы РНК получать комплементарные фрагменты ДНК. Напомним, что при синтезе белка с гена считывается информация в направлении ДНК —> РНК. При обратной транскрипции процесс идет в противоположном направлении.
Подлинную революцию в изучении генома совершило открытие в 1983 г. полимеразной цепной реакции (ПЦР). Ее автору, К. Муллису в 1993 г. была присуждена Нобелевская премия в области биологии и медицины. Метод ПЦР позволяет тиражировать in vitro фрагменты ДНК в неограниченном количестве, при этом в качестве исходного материала достаточно минимального количества ДНК. Процесс амплификации ДНК состоит из повторяющихся циклов, каждый из которых включает три стадии: денатурации (получение одноцепочечных фрагментов ДНК), отжига (присоединение праймеров к комплементарным последовательностям одноцепочечных молекул) и синтеза.
Эти и некоторые другие методы генетической инженерии лежат в основе молекулярно-генетической диагностики. Коротко остановимся на некоторых этапах молекулярно-генетических исследований.
Для начала необходимо получить образцы ДНК для анализа. Исходным материалом для получения ДНК могут служить любые клетки организма, содержащие ядро. Чаще всего используют лейкоциты периферической крови. Для одного анализа необходимо от нескольких нанограммов до нескольких микрограммов ДНК. Для получения такого количества ДНК потребуется не более 1 мл крови. Для некоторых анализов достаточно всего одной капли крови. Вся ДНК клетки называется геномной ДНК, поскольку содержит весь геном. Часто для анализа требуются не вся ДНК, а лишь определенные ее фрагменты, в которых расположены изучаемые гены.
Для проведения анализа необходимо иметь достаточное количество ДНК. Эта задача сейчас решается с помощью описанной выше полимеразной цепной реакции (ПЦР).
Необходимым этапом является также рестрикция (разрезание) ДНК под действием специальных ферментов (рестриктаз), которые способны распознавать специфические короткие (4-10 пар нуклеотидов) последовательности. Каждая рестриктаза разрезает цепь ДНК в строго определенном месте. В результате действия каждой рестриктазы образуется специфичный для нее набор фрагментов различной длины.
Далее смесь фрагментов подвергают электрофорезу в специальном геле. В зависимости от своей длины фрагменты движутся в геле с различной скоростью. По окончании электрофореза фрагменты ДНК занимают каждый определенное положение в виде полосы, однако, поскольку размер генома велик, полосы после электрофореза расположены настолько тесно, что их невозможно различить.
Чтобы выявить специфические фрагменты ДНК, проводят еще одну процедуру, которая называется блот-гибридизацией по Саузерну. Эта методика состоит из нескольких этапов и приводит к получению четкой картины фрагментов ДНК, напоминающей штрих-код.
Для обнаружения конкретных мутаций проводят секвенирование ДНК, т.е. определение точной последовательности пар нуклеотидов в ДНК. Существуют различные модификации описанных методов, однако главной целью работы в любом случае является получение максимально возможной информации о строении гена.
Моделирование на животных
Преимущества генетических исследований на животных очевидны - это широкие возможности для скрещивания и отбора, контроля условий среды, в которых развивается потомство, отсутствие этических преград для проведения любых манипуляций на всех уровнях функционирования, начиная с геномного и кончая популяционным. Спектр возможностей исследований человека гораздо уже. Поэтому психогенетика часто использует методы моделирования на животных различных состояний, некоторых психических заболеваний, элементарных форм научения.
У человека и других млекопитающих гораздо больше общности, чем различий. Например, число нуклеотидов в ДНК человека и мыши составляет примерно три миллиарда, причем более 99% генов у них являются общими. Любое поведение является результатом деятельности мозга. Известно, что многие механизмы деятельности мозга у человека и животных также идентичны. Поэтому нет ничего удивительного в том, что эксперименты на животных, как на более простых системах, позволяющих к тому же осуществлять вмешательство в деятельность этих систем, позволяют подойти к решению некоторых проблем генетики поведения человека. Вообще генетика поведения животных является самостоятельной областью генетики поведения, и в данной области ведутся различные исследования, прямым образом не связанные с генетикой человека. Мы не будем подробно касаться этой сферы, а упомянем лишь несколько примеров исследований на животных, которые способствуют пониманию проблем генетики поведения человека. В дальнейшем, особенно в том разделе, где речь пойдет об индивидуальном развитии, мы еще не раз столкнемся с модельными экспериментами, ведущимися на животных. Воспользуемся несколькими примерами модельных экспериментов на животных.
Способность к обучению.
Никто не станет отрицать, что способность к обучению присуща не только человеку, но и животным. Эта способность широко исследуется в генетике поведения животных. Исследования проводятся также в основном на инбредных линиях крыс или мышей. В этой области преимущество экспериментов на животных заключается в том, что по результатам скрещивания особей разных линий, т.е. животных с разными "поведенческими" генами, можно составить представление о характере наследования способности к обучению, а возможность выращивать потомство в различных условиях среды помогает определить характер средовых влияний на формирование способности к обучению, что в экспериментах с человеком, естественно, невозможно.
В ряде экспериментов изучались различия между линейными животными по обучению условной реакции избегания. Животных помещали в специальную клетку, разделенную на два отсека. В один из отсеков подавался электрический ток, вызывающий неприятные ощущения. Чтобы избежать неприятного воздействия, животное должно было перепрыгнуть в другой, безопасный отсек клетки. Сначала животное обучается этой реакции, а затем перед включением тока подается сигнал в виде вспышки света, и животное постепенно обучается реагировать на включение вспышки, а не на сам ток. Показателем скорости обучения служит число вспышек, необходимое для формирования условной реакции избегания. Оказалось, что инбредные линии мышей значительно отличаются по скорости обучения. Скрещивание линий с высокой и низкой скоростью обучения показало, что в основе различий лежит простой моногенный тип наследования.
Другой тип поведения, связанного с обучением, - обучение в лабиринте - в отличие от поведения избегания, оказался подвержен не только генетическим, но и средовым влияниям. Чтобы добраться до кормушки, крысам необходимо было преодолеть лабиринт. В исходной популяции встречались животные, которые быстро находят путь и делают мало ошибок, и, наоборот, такие, которые затрачивают на это много времени и часто ошибаются. Выборочная селекция в нескольких поколениях позволила выделить неперекрывающиеся субпопуляции "умных" и "глупых" животных. Это говорит о том, что в исходной популяции существовала заметная генетическая изменчивость по способности к обучению в лабиринте.
Затем был осуществлен эксперимент, который чрезвычайно важен для формулировки гипотезы в отношении способности к обучению у человека. Детенышей каждой из линий - "умной" и "глупой" - разделили на 3 группы. Крысята первой группы выращивались в обычных условиях. Детеныши из второй группы с момента рождения были лишены тех факторов среды, которые способствуют познавательной или исследовательской деятельности, т.е. росли в условиях депривации. Третья группа крысят, наоборот, была помещена в обогащенную среду - внутри клетки находились трапы, зеркала, туннели и другие приспособления, способствующие развитию разнообразных видов активности. Самое интересное заключается в том, что межлинейные различия в этой ситуации наблюдались только в группах животных, выращиваемых в обычных условиях. Депривированные животные обеих линий одинаково плохо справлялись с лабиринтом, а вот животные, росшие в сенсорно-обогащенной среде, независимо от принадлежности к линии, почти одинаково хорошо обучались поведению в лабиринте.
Из этого опыта следует, что депривация в раннем возрасте отрицательно сказывается на способности к обучению в дальнейшем, и, наоборот, сенсорно-обогащенная среда может улучшить эту способность, несмотря на существующие генетические различия.
Подчеркнем еще раз, что генетические различия, заметные в одном диапазоне сред, могут не проявиться в фенотипе при перемещении тех же генотипов в другой диапазон, и что генетическая детерминации вариативности признака не означает невозможности его изменения путем изменения среды. Здесь же хочется обратить внимание читателей на гипотетический график нормы реакции для интеллекта человека (рис. 8.11), демонстрирующий критическое значение депривации для развития нормального интеллекта. Так что между генетическими исследованиями поведения животных и человека существует множество точек соприкосновения, и некоторые схемы эксперимента, которые мы можем осуществить на животных, позволяют проверять наши гипотезы в отношении человека.
Алкоголизм.
При изучении наследственных причин алкоголизма у человека широко используются эксперименты на мышах и крысах. Исследователям удалось путем инбридинга (близкородственного скрещивания) получить чистые линии мышей с различной потребностью в алкоголе (напомним, что все животные, принадлежащие к одной линии, имеют почти одинаковые генотипы). Мыши некоторых линий полностью избегают алкоголя, тогда как другие предпочитают в среднем до 2/3 дневной нормы жидкости получать в виде 10% раствора алкоголя. Были исследованы межлинейные различия в свойствах мозга и печени животных. Основные различия между животными, принадлежащими к контрастным линиям, выявились в двух сферах - в метаболизме алкоголя и в его воздействии на физиологию мозга. Было показано, что повышенная потребность в алкоголе сочетается с высокой чувствительностью мозга к его наркотическому действию.
Эксперименты на животных позволили очертить те области, в которых необходимо было искать генетические различия между людьми - это метаболизм алкоголя, т.е. комплекс его превращений в организме, и нейрофизиологические реакции на алкоголь. В исследованиях на близнецах было обнаружено существенное генетическое влияние на метаболизм алкоголя, а в нейрофизиологических экспериментах с регистрацией электроэнцефалограммы (ЭЭГ) было показано, что лица, склонные и не склонные к потреблению алкоголя, имеют разный характер ЭЭГ и разную ее реактивность на введение алкоголя. Оказалось, что родственники алкоголиков, не страдающие алкоголизмом, имели похожий характер биоэлектрической активности мозга, а МЗ близнецы, в отличие от ДЗ, демонстрировали высокое сходство по реактивности ЭЭГ на алкоголь. Все эти эксперименты позволили заключить, что некоторые особенности метаболизма и характер чувствительности нервной системы к алкоголю являются генетически обусловленными причинами различий между людьми в склонности к потреблению алкоголя. Возникновение же алкоголизма как болезни, со всем комплексом поведенческих и личностных расстройств, определяется в основном внешними обстоятельствами, т.е. средой. В обществе, где алкоголь доступен и традиционно является одним из элементов среды при общении людей, генетически восприимчивый к алкоголю человек имеет больше шансов заболеть алкоголизмом.
Помимо способности к обучению и алкоголизма на животных моделируются агрессивность, склонность к беспокойству и страху, а также такие заболевания человека, как эпилепсия и болезнь Альцгеймера.
С развитием генной инженерии стали появляться принципиально новые возможности исследований эффектов единичных генов. В последние годы все чаще исследуются так называемые трансгенные мыши и мыши-нокауты. Разработаны методы внедрения в геном чужеродного гена (трансгенные мыши) и выключения (нокаута) одного из естественных генов. При нокауте один из нормальных генов полностью инактивирован. Нокаут напоминает прием, используемый в нейрофизиологии для изучения последствий удаления отдельных областей мозга. В случае генного нокаута изучаются последствия удаления отдельного гена, т.е. фактически последствия лишения организма того генного продукта, который вырабатывается на основе изучаемого гена.
Оба метода сложны в осуществлении. Работа ведется в несколько этапов, которые включают клонирование нужного гена в бактериях, работу с генной ДНК, внедрение измененной ДНК в эмбриональные клетки на ранних стадиях развития, различные ухищрения для отбора нужных клеток, в которых произошла инактивация гена или, наоборот, был внедрен "чужой" ген, последовательные скрещивания для получения поколения мышей с нужным геномом. В дальнейшем это потомство тщательно исследуется для выявления морфологических, физиологических и поведенческих нарушений, являющихся последствиями вмешательства. К сожалению, такие методически сложные процедуры нередко заканчиваются лишь констатацией наличия плейотропного эффекта действия гена. Плейотропный, или множественный, эффект означает, что генный продукт может быть включен в столь важные процессы метаболизма, что они существенно влияют на самые разнообразные признаки, в том числе и на поведение. Однако уже сейчас получены первые результаты, демонстрирующие, что утеря некоторых важных биохимических субстанций в нервной клетке в связи с нокаутом или внедрение новых генов могут вызывать специфические изменения поведения.
Еще один интересный факт заслуживает внимания. Не так давно исследователи генетики поведения животных провели своеобразные "эксперименты-близнецы". В трех независимых лабораториях (в штатах Орегон и Нью Йорк в США и в штате Альберта в Канаде) были выполнены идентичные исследования. В каждой из лабораторий ученые постарались максимально уравнять все генетические и средовые переменные: выбрали генетически идентичные линии животных (мышей), применили одну и ту же схему эксперимента, максимально контролировали все условия среды (питание животных, световой режим, время проведения тестирования и т.п.). Однако, несмотря на это, ни в одной из лабораторий не удалось получить полностью совпадающих результатов. Этот факт свидетельствует о том, что существует множество средовых эффектов и случайностей, которые трудно уравнять, например, запахи в лабораториях. Среда оказывается настолько вариабельной, что не поддается полному контролю. Этот эксперимент продемонстрировал, что даже незначительные различия в среде могут приводить к заметным изменениям в поведении. Тем более важно это учитывать в исследованиях, проводимых на человеке.
Выводы
Геномика и психогенетика
Геномом называется полный состав ДНК клетки.
Геномика - это молодая интенсивно развивающаяся отрасль генетики, изучающая принципы построения геномов и их структурно-функциональную организацию.
Структурная геномика изучает нуклеотидные последовательности ДНК, в том числе строение и локализацию генов. Одной из задач структурной геномики является построение генетических карт организмов.
Функциональная геномика решает задачи идентификации функций отдельных участков генома и механизмы их взаимодействий в клеточном ансамбле.
Современные представления о геноме человека базируются на нескольких открытиях молекулярной биологии 70-х годов ХХ века, основными из которых являются открытие полимеразной цепной реакции (ПЦР), позволяющей получать достаточное количество ДНК для анализа, и разработка методов секвенирования, которые позволяют расшифровывать точную последовательность нуклеотидов в цепях ДНК.
В конце 80-х годов ХХ столетия началось осуществление международного проекта "Геном человека", основной задачей которого было секвенирование генома человека. Полное секвенирование генома человека было завершено в 2000 году.
Секвенирование генома человека привело к открытию огромного количества однонуклеотидных полиморфизмов (ОНП) - генетических вариантов последовательностей нуклеотидов одного и того же участка ДНК у разных людей. Распределенные по всему геному ОНП используются в качестве генетических маркеров.
По расчетам генетиков люди оказываются идентичными по нуклеотидным последовательностям на 99,9%. Таким образом, генетическая вариативность людей возникает за счет различий лишь в 0,1% генома.
Данные секвенирования показали, что в геноме человека чуть более 30000 генов. Большинство генов имеют размеры до 50000 пар нуклеотидов. Число синтезируемых белковых продуктов превышает число генов в 1,5-2 раза. Это является результатом альтернативного сплайсинга.
В современной психогенетике используются три основных подхода к изучению поведенческой геномики: анализ сцепления, анализ ассоциаций, непосредственный анализ ДНК (секвенирование и идентификация мутаций).
Анализ сцепления
Классический вариант анализа сцепления использован на использовании явления кроссинговера. В процессе кроссинговера гены, расположенные в одной хромосоме (сцепленные), рекомбинируют тем чаще, чем дальше они отстоят друг от друга. Т.Морган в начале ХХ века предложил использовать явление сцепления для создания генетических карт, т.е. схем расположения конкретных генов на конкретных хромосомах.
Для локализации гена на хромосоме необходим маркер, т.е. ген, локализация которого известна, либо морфологический или молекулярный маркер.
При анализе сцепления мерой "генетического расстояния" служит частота рекомбинации между маркером и исследуемым геном. Расстояние между генами измеряется в сантиморганидах (сМ). Одна сантиморганида соответствует расстоянию между генами, рекомбинация которых происходит с частотой 1%. При частоте рекомбинации 50% считается, что гены наследуются независимо.
Классический анализ сцепления чаще всего используется для локализации генов альтернативных (качественных) признаков. Для количественных признаков используется несколько иная модель картирования.
Локусы количественных признаков (ЛКП) - это полигенные системы, обеспечивающие непрерывную вариативность признака в популяции.
Метод картирования ЛКП основан на исследовании ДНК у пар близких родственников (чаще всего сибсов). Идея метода состоит в том, что любой ребенок в семье может иметь от 0 до 2 общих аллелей с любым из своих братьев и сестер. Если разница в количестве совпадающих аллелей никак не сказывается на количественных соотношениях признака у сибсов, значит тестируемый ген не имеет отношения к признаку. Соответственно, обратная картина говорит о причастности гена к детерминации изучаемого признака.
Целью картирования ЛКП является поиск не единичного гена, а, скорее, групп генов, принимающих участие в формировании популяционной вариативности по количественному признаку. Метод картирования ЛКП эффективен для выявления эффектов главных генов, вносящих основной вклад в формирование количественного признака.
Анализ ассоциаций. Метод гена-кандидата
Метод анализа ассоциаций по сравнению с обычным анализом сцепления, может обнаруживать ЛКП, которые отвечают за меньший процент вариативности. Ассоциация - это корреляция между отдельными аллелями и признаками в популяции.
В исследованиях, использующих метод ассоциаций. сначала формируется гипотеза о связи какого-либо гена с признаком или заболеванием. Такой ген носит название гена-кандидата.
Гены, кодирующие белки, участвующие в процессах передачи информации в нервной системе, привлекают внимание исследователей как возможные гены-кандидаты для поведенческих признаков (например, гены кодирующие информацию для синтеза серотонина или дофамина).
В ассоциативных исследованиях сопоставляется встречаемость определенного аллеля гена-кандидата в популяции в целом и у лиц, обладающих интересующим признаком или пораженных болезнью. Если данный аллель чаще встречается у носителей более выраженного признака или заболевания, можно предположить участие этого аллеля в наследственной детерминации данного признака.
Прямой анализ ДНК и выявление мутаций
Определение местоположения гена на хромосоме не является основной целью молекулярных исследований в психогенетике. Важнее знать, не где расположен ген, а как он работает и как факторы среды влияют на его активность и формирование изучаемой поведенческой характеристики.
В основе молекулярно-генетического анализа лежат методы генной инженерии.
Основные этапы прямого анализа ДНК включают:
получение образцов ДНК;
амплификацию ДНК с помощью ПЦР;
рестрикцию (разрезание) ДНК на фрагменты;
электрофорез смеси фрагментов ДНК, позволяющий дифференцировать фрагменты различной длины;
блот-гибридизацию фрагментов ДНК, позволяющую получить схему (своеобразный штрих-код) участков ДНК, идентичных для конкретных родственников;
секвенирование ДНК (определение конкретной последовательности пар нуклеотидов) изучаемого отрезка ДНК.
Моделирование на животных
Генетические исследования животных, в отличие от человека, обладают более широким спектром возможностей (скрещивания, селекция, манипуляции со средой, возможность вмешательства в функционирование организма для изучения внутренних механизмов и т.п.).
Благодаря значительной общности в организации геномов млекопитающих и человека, а также сопоставимости функционирования их нервной системы, в психогенетике используются методы моделирования на животных различных психических состояний, заболеваний, элементарных форм научения и др.
Модельные эксперименты на животных широко используются при изучении наследственности алкоголизма, эпилепсии, болезни Альцгеймера, агрессивности, склонности к беспокойству и страху, способности к научению.
Материал взят из электронного учебника «Психогенетика» с сайта http://www.ido.edu.ru
в начало
|
|
|
|